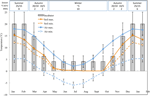
Obrázek 1. Měsíční maximální a minimální údaje o teplotě vzduchu a půdy v australských Alpách (průměr ± STDEV). Teploty vzduchu jsou průměry z databáze bomb, teploty půdy byly měřeny pomocí ibuttonů (viz metody). Tyče označují experimentální teploty inkubátoru (průměr ± STDEV)., Ekvivalentní roční období (sezóna), teplotní režim inkubátoru (°C, 12/12 h, světlo / tma) a doba trvání léčby (týdny), jak je uvedeno v horním panelu.
S cílem odhalení teplotních podmínek blízkých těm, které semena zkušenosti post-rozptýlení, denní maximální a minimální teploty půdy byly údaje shromážděny v rámci studie stránky (Seaman ‚ s Hut; 36.27°S a 148.17°E, 2030 m.s.l.) pomocí ibutton záznamníky dat (n = 10, Vložené Data Systems, USA) umístil na 4 cm pod povrch půdy na základně vegetace od 17. ledna do 17. prosince 2012., Umístili jsme dřevorubci na 4 cm, aby se zabránilo narušení povrchu, ale stále být reprezentativní pro podmínky na kterém jsou semena vystavena v půdě; tato hloubka je střední, co byl použit v předchozí alpské půdy sledování teploty cvičení (Scherrer a Körner, 2011; Pauli et al., 2015). Podle podezření průměrná teplota půdy neklesla během zimy pod bod mrazu (Obrázek 1). V létě je známo, že teplota půdy pod vegetací sleduje okolní teplotu, zatímco teploty holé půdy překročí okolní teplotu až o 15°C (služba ochrany půdy nezveřejněné záznamy, 1960–1970).,
sběr a klíčení semen
zralá semena 54 druhů ze 16 rodin a 37 rodů byla shromážděna mezi lednem a dubnem 2009, 2010 a 2011 (úplné názvy a úřady viz tabulka 1. Poukázky byly podány v australském Národním herbáři Canberra). Celkem druhů představují více než čtvrtinu Australského krytosemenných rostlin nalezené v alpských regionech (Costin et al., 2000, viz tabulka 1), ačkoli mnoho z těchto druhů se rozšiřuje i pod treeline., Životaschopnost všech sbírek byla odhadnuta před výsevem v experimentálních podmínkách klíčení pomocí barvicí techniky tetrazoliumchlorid (TZ) (International Seed Testing Association, 2003). Další podrobnosti o sběru a zpracování viz dodatek A v doplňkovém materiálu.
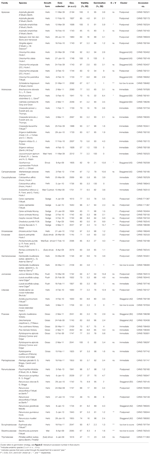
Tabulka 1. Studijní druhy, rodina a autorita, forma růstu a podrobnosti o sbírkách, životaschopnost procentního osiva (TZ-odhadovaná, ± s. e.), konečné procento klíčivosti dosažené (± s. e.,), statistický rozdíl mezi životaschopností a klíčivostí (g < v, testováno pouze v případech, kdy klíčení bylo menší než životaschopnost).
Fenologie klíčení semen byl zkoumán napodobováním teplotní režimy, které alpine semena zkušeností in situ, post-rozptýlení, v uměle zkrácený průběh ročních období. Semena byla v průběhu experimentu napuštěna, takže výsledky naznačují potenciál klíčení,když voda neomezuje., Vzduchu a teplota půdy údajů (viz Obrázek 1) byly použity jako vodítko a ověřit inkubátor teplotní režimy, i když logistickou contraints práce s inkubátory brání začlenění výkyvy vlastní přírodní podmínky. Testy klíčení použily osm replikátů 25 semen na sbírku, zasetých do plastových Petriho nádob o průměru 9 cm obsahujících 1% čisté vody-agar. Petriho nádobí bylo utěsněno parafilmem, aby se zabránilo vysychání agaru, než bylo umístěno do inkubátorů klíčení (Thermoline Scientific, Melbourne, NSW, Austrálie)., Polovina nádobí byla zabalena do hliníkové fólie, aby se vyloučilo světlo a byly uvedeny požadavky na světlo pro klíčení. Každá ze čtyř replikátů na ošetření světla na sbírku byla umístěna na jiné polici ve stejném inkubátoru a umístění v této polici bylo znovu uspořádáno týdně. Všechny replikáty každého sbírky byly přesunuty prostřednictvím následujících simulované thermoperiods: 8 týdnů při 25/15°C (den/noc teplota) → 2 týdny na 20/10°C → 2 týdny na 10/5°C → 10 týdnů v konstantní 4-5°C → 2 týdny na 10/5°C → 2 týdny na 20/10°C → 6 týdnů při 25/15°C., Jeden cyklus celého experimentu tak napodoboval teploty připomínající léto, začátek podzimu, pozdní podzim, zimu, brzy na jaře, pozdní jaro a léto a trval celkem 32 týdnů. Zkrácený rok byl považován za rozumný, protože přistoupení obecně ukončilo další klíčení v dané léčbě před přesunem do další „sezóny“., 12 / 12 h světlo/tma fotoperioda byla poskytnuta v celém tím, zářivky (> 90 µmol m−2 s−1 ve středu každé police), a na jaře, v létě a na podzim režimy teploty světla se shodoval s teplota teplé období dne. Klíčení, definované jako kořínek vznik více než 1 mm, byl skóroval každých 7 dní, a vyklíčila semena a sazenice byly následně odstraněny z nádobí. Klíčení fólií zabalených desek bylo hodnoceno jako výše, ale pod velmi nízkým zeleným světlem.,
po jednom experimentálním cyklu (32 týdnů) byla většina sbírek ukončena (až na výjimky viz doplňující informace), přičemž v tomto okamžiku vykazovala nulové klíčení po dobu nejméně 2 týdnů. Po ukončení byly všechny zbývající neporušené semena rozřezány skalpelem pod mikroskopem. Semena s pevným, čerstvým endospermem a embryem byla považována za životaschopná a semena prázdná z embrya byla odečtena z celkového počtu při výpočtu procentního klíčení pro každý odběr., Vypočítali jsme index, světla požadavky na klíčení pro jednotlivé druhy jako zlomek klíčení ve tmě ošetření v době, která kumulativní klíčení percentil 50 a 95% bylo dosaženo ve střídání světlo/tma léčby (viz níže).
Ekologické Koreláty Klíčení Strategie
Jsme sestavili databázi druhů funkční vlastnosti a geografické opatření z publikovaných údajů pro posouzení korelace s klíčení strategie., Databáze zahrnovala následující nepřetržité rysy: průměrná hmotnost osiva, požadavek na světlo pro klíčení (viz výše), specifická plocha listů (čerstvá Plocha/suchá hmota), individuální plocha listů, individuální suchá hmotnost listů a výška při zralosti. Minimální, průměrné a maximální sběr nadmořských výškách pro druhy byly stanoveny ze všech herbář záznamy v Australském Virtuální Herbář, a nadmořská výška, v níž konkrétní seed kolekce, použitý ve studii byl proveden byl také jako kontinuální vlastnosti. Forma růstu byla zaznamenána jako jedna ze tří kategorií: graminoid, bylina nebo keř., Zahrnuli jsme také binární rys: zda semena byla endospermická nebo ne.
vlastnost údaje byly shromážděny z různých zdrojů, včetně Australské Virtuální Herbář (http://avh.chah.org.au/), Australské Národní Herbář vzor informace rejstřík (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Alpské Flóry (Costin et al., 2000), Flora Nového Jižního Walesu (Harden, 1990) a srovnávací vnitřní morfologie semen (Martin, 1946). Hmotnost osiva a specifická plocha listů byly získány z jednotlivých terénních sbírek používaných ve studiích klíčení., Hmotnost osiva byla získána vážením čtyř šarží 10 semen. SLA byla stanovena na základě pěti plně rozšířených listů od jednotlivých jedinců na druh. Ty byly shromážděny v oboru, skenování na plochém skeneru, suší se při 60°C do konstantní hmotnosti a zváží na nejbližší 0.000 g. Kde nedostatečné semeno bylo k dispozici, nebo specifické listové plochy nejsou k dispozici, údaje byly získány z jiných ANBG sbírek stejného druhu nebo z jiných zdrojů dat (S. Venn, nepublikované údaje)., Zda bylo nebo nebylo osivo endospermické, bylo stanoveno z výkresů osiva v záznamech banky ANGB Seed bank po konzultaci se zaměstnanci banky ANBG seed. Kompletní databáze a zdroje jsou k dispozici na vyžádání od autorů.
Analýza Dat
Klíčení vzory byly hodnoceny vizuálně a pomocí shlukové analýzy v PC-ORD pro Windows, verze 6 (MJM Softwarový Design, Gleneden Beach, OR, USA). Analýza použila euklidovskou matici vzdálenosti a algoritmus spojování nejbližšího souseda., Datové body pro analýzu clusteru byly čas (týdny) na specifické kumulativní percentily klíčení (t20, t40, t50, t60, t80 a t95%) pro každý sběr (průměrně přes 4 nádobí). Tedy kolekci, která vyklíčila brzy v experimentu by mít krátkou dobu na percentily až t80% například, vzhledem k tomu, že kolekce, která odloženo klíčení, dokud se teploty zvýšily v návaznosti na simulované zimě by mít dlouhé časy, aby tyto percentily., Složitější klíčení vzory, jako jsou pomalý, stabilní klíčení nebo výbuchy klíčení, by také mohl být detekován, a by být indikována rovnoměrně rozložených časech nebo obdobích, seskupený na začátku a na konci experimentu, resp. Tato metoda byla použita jako způsob, jak uložit objektivní posouzení klíčení vzoru, nezávisle na teplotě sama o sobě.,
jednosměrná analýza rozptylu (ANOVA) byla provedena pro replikaci desky každé kolekce posoudit, zda existuje významný rozdíl mezi procento životaschopnosti (jak stanoví TZ test), a konečné procento klíčení. Další obecné lineární modely byly použity k posouzení, zda se klastry vyplývající z analýzy klastru liší životaschopností nebo konečným procentem klíčení.
asociace mezi ekologickými vlastnostmi a klíčivými strategickými klastry byly testovány dvěma způsoby., Za prvé, jsme testovány na mysli rozdíly mezi shluky pomocí ANOVA s clusteru jako fixní efekt a ekologické koreláty jako proměnné odezvy, nebo pomocí replikované G-testy pro endospermy a růst podobě. Za druhé, protože klíčení a další funkční rysy jsou vysoce pravděpodobné, že budou ovlivněny evoluční historií, hodnotili jsme tyto korelace pomocí fylogenetických regresí. Fylogeneze byla konstruována pro studijní druhy s použitím Phylocom Phylomatic a bladj balení v R (Webb et al., 2008; R Core Team, 2013)., Hodnotili jsme přítomnost fylogenetické struktury v proměnných pomocí fylosigu nebo phylo.d (diskrétní nebo binární proměnné) funkce balíčku Fytoolů v R (Revell, 2012). Fylogenetické regrese byly prováděny pomocí funkce pgls v Caperu (Orme et al., 2013). Analýzy byly prováděny strukturovaným způsobem odrážejícím shlukování odhalené v PC-Ord (viz níže)., Zpočátku jsme zařadili čtyři klíčení strategie (odloženo, bezprostřední, a potácel kategorií v rámci každé předchozí), pak tři (házet do jednoho pytle všechny rozložené druhů a v porovnání s odloženo a intermediate) a nakonec jsme zhodnotili rozdíly mezi dvěma nejširší strategie, odloženo a středně pokročilé.
výsledky
životaschopnost osiva ve srovnání s konečným klíčením
celková životaschopnost osiva a konečné procento klíčivosti byly vysoké. Průměrná TZ-odhadovaná životaschopnost ve všech sbírkách byla 80 ± 1.,7%, přičemž více než dvě třetiny sbírek vykazují více než 75% životaschopnost (Tabulka 1). Tam byl žádný rozdíl v kvalitě sbírek po celé 3 roky (průměr rentability v letech 2009, 2010 a 2011 bylo 79 ± 2.8, 80 ± 2.7 a 81 ± 3.7%), což naznačuje, že bankovní semen sebraných v roce 2009 se nesnižuje jejich životaschopnost.
klíčení, které překročilo nebo nebylo významně nižší než TZ-odhadovaná životaschopnost byla dosažena u 45 z 54 druhů (jednosměrná ANOVA: p>0.05, Tabulka 1)., Ze zbývajících devíti druhů bylo konečné klíčení čtyř druhů velmi nízké (< 5%), navzdory dobré životaschopnosti (ca. 55-94%), a tak jsme nebyli schopni vyvodit žádné závěry o strategiích klíčení těchto druhů. Neschopnost klíčit by mohla odrážet buď to, že naše léčba nezmírnila dormancy, nebo že semena nebyla při sběru plně zralá. Řez testy na konci experimentu ukázaly, že ve všech případech, většina z osn-naklíčených semen obsahovala zdravé, do sebe vstřebala embryo., Zajímavé je, že 34 druhů dosáhlo klíčení, které bylo větší než odhadovaná životaschopnost TZ (Tabulka 1), což naznačuje, že test TZ běžně podceňoval životaschopnost osiva.
klastry klíčení
analýza clusteru odhalila dva hlavní klastry (A A B), s podstrukturou v každém (Obrázek 2). Pro většinu druhů (n = 21 druhů z 10 rodin), klíčivost byla odložena během zimních teplotách (5°C den/noc) nebo dokud teplota byla zvýšena na 10/5°C po zimě (A1 a A1:zimní, Čísla 2, 3A)., Alternativně klíčení začalo okamžitě při 25/15°c (n = 17 druhů, šest rodin, vidět v clusteru B1, obrázky 2, 3B). U většiny těchto druhů byla rychlost klíčení rychlá; 50% celkového klíčení (t50) se vyskytlo za méně než 3 týdny (Doplňkový Obrázek 1).
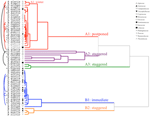
Obrázek 2. Cluster analýza dendrogram na základě času na kumulativní klíčivost percentilů (20, 40, 50, 60, 80, A 95%). Druh označený rodem počáteční a první část názvu druhu. Hvězdičky označují sbírky, které byly před testováním nakloněny., Jemné oblouky spojují duplicitní sbírky a porovnávají vliv bankovnictví na klíčení. Symboly označují různé rodiny (viz legenda). Sbírky zobrazené v šedém textu jsou ty, které byly vystaveny dvěma cyklům experimentu.
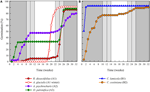
Obrázek 3. Kumulativní procentuální klíčení jednoho reprezentativního druhu z každé klíčivosti. A) shluk a, druhy, které odložily klíčení, B) shluk B, druhy, které okamžitě vyklíčily., Všimněte si, že rozložené vzory klíčení jsou vnořeny do výše uvedených klastrů. Teplotní režimy inkubátoru jsou reprezentovány stínováním (viz Obrázek 1).
v každé z těchto širokých tříd však existovaly jasné dílčí klastry odrážející klíčení, které bylo v průběhu času rozloženo, tj. Potácel klíčení došlo ve 13 druhů z šesti rodin a došlo především, když denní teploty překročily 10°C (Obrázky 3A,B)., Tyto rozložené germinátory byly v analýze clusteru rozděleny do tří skupin. Cluster A2 se sedmi sbírkami vykazoval značné klíčení jak před, tak po období studené teploty. Kde klíčení přestal před teplotách měněn jsme dospěli k závěru, že nedostatek další pre-zimní klíčení nebyla řízena teplota (A3). Naopak klíčení velké části skupiny A2 se zdálo být zastaveno pouze tehdy, když byly teploty sníženy na 10/5°C., Cluster B2 obsahoval další čtyři druhy, které vykazovaly podstatné klíčení brzy v experimentu a šel k dosažení relativně malý podíl z jejich celkových klíčení po teploty byly vráceny do výše 5°C (Obrázek 3B).
ve srovnání s relativně malou variací mezi druhy v klastrech A1 a B1 vykazují rozložené klastry také mnohem větší diferenciaci mezi druhy. Jednotlivé křivky pro všechny druhy jsou uvedeny na doplňkových obrázcích 1i-VII. podrobnější informace o strategii klíčení viz dodatek B v doplňkovém materiálu.,
Ekologické Koreláty Klíčení Strategie
všechny ekologické koreláty, že jsme uvažovali pouze ty přímo spojené s semen vlastnosti byly významně koreluje s klíčení strategie bez ohledu na to, zda jsme hodnotili korelaci na úrovni dvou hlavních klastrů (odloženo vs. okamžité klíčení), nebo na úrovni tří nebo čtyř klastrů, nebo zda jsme tvořily fylogenetická struktura, nebo ne (Tabulka 2)., Druhy s odloženým klíčením měly těžší semena a byly pravděpodobnější, že budou endospermické ve srovnání s druhy, které okamžitě vyklíčily (obrázek 4). Druhy s rozloženým klíčením byly Střední v osivové hmotě a endospermii. Ne všechny charakteristiky osiva však korelovaly se strategií klíčení: neexistovala žádná korelace mezi lehkými požadavky na klíčení semen a strategií. V rozporu s naším očekáváním, druhy s vyšší specifickou listovou plochu (mohly naznačovat vyšší míru růstu) byly více pravděpodobné, že vykazují okamžité klíčení., Podobně u druhů s vyšší průměrnou nadmořskou výškou nebo menšími nadmořskými výškami nebyla větší pravděpodobnost odloženého klíčení.

Tabulka 2. Výskyt fylogenetické struktury v, a koreluje, strategie klíčení.
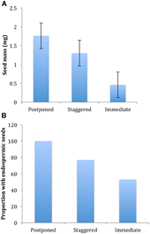
Obrázek 4., Rozdíly v (A) průměrná hmotnost semen a (B) podíl druhů s endospermic semen mezi druhy s odloženo (cluster A1), rozložené (kombinace clusteru A2, A3 a B2) a okamžitou klíčivost (cluster B1) strategie. Rozdíly mezi strategiemi jsou významné při p ≤ 0,05, viz tabulka 2.
Diskuse
Klíčení ve vysokohorských stanovišť byla v minulosti považována za ekologicky kontrolované, s zimě sněhu izolační semena proti potenciální klíčení podněty a vykreslování dormance zbytečné (Billings a Mooney, 1968)., Naše výsledky však podporují novější důkazy o tom, že spící mechanismy jsou běžné u alpských druhů jak v Austrálii (Hoyle et al., 2013; Sommerville et al., 2013) a jinde po celém světě (Cavieres and Arroyo, 2000; Schutz, 2002; Shimono and Kudo, 2005; Mondoni et al., 2009; Schwienbacher et al., 2011)., Dormance se zdá hrát významnou roli při kontrole in situ načasování klíčení mnoha Australské alpské druhy, působící na zpoždění, odložit, nebo zpomalit rychlost klíčení a tedy potenciálně svěřují riziko-odmítavý regenerace strategie v drsném a proměnné prostředí. Snad nejvýraznější z našich klíčení výsledky byl podíl druhů, pro které klíčení strategie měnit v sběr semen pro daný druh, což naznačuje, že oba spící a non-spící vlastnosti byly vystaveny v rámci stejné sběr semen těchto druhů., Zejména strategie klíčení byly obecně velmi zachovány v rodinách a také korelovaly s prvky anatomie semen: hmotnost a endospermie. Strategie klíčení však byly nezávislé na jiných ekologicky významných funkčních vlastnostech zralé rostliny.
Rozdíly v Klíčení Strategie
Semena téměř polovina studovaných druhů se objevila schopny klíčit, dokud byli vystaveni chladné, vlhké období (konstantní 5°C), což naznačuje, že studená stratifikace zmírnit fyziologické dormance mechanismus., Odložení klíčení až do následujícího jara může semenáčkům umožnit vyhnout se nastolení přes nebo před krutou zimou a zároveň optimalizovat krátkou nadcházející vegetační sezónu. Na rozdíl od oportunistické klíčení vystavoval druhů naklíčených ihned mohou poskytovat selektivní výhodu při riziku zimní sazenice úmrtnost je nízká, tím, že umožňuje rostliny na květ dříve, následující jaro nebo ve větší velikosti (Donohue, 2002).,
dosud nebyly publikovány žádné důkazy o intra-specifické variabilitě mechanismů nečinnosti v rámci sbírek alpských semen, jako by to vysvětlovalo rozložené strategie klíčení zde pozorované. Klíčení rozložené v průběhu času, může být také vysvětlen různou úrovní zralosti semen mezi jednotlivci v populaci v době odběru, a/nebo by mohlo odrážet dimorfní strategie v rámci jedné rostliny spojené s postavení na rostlině nebo načasování vývoje.,
Evoluční bet-hedging je často vyvolán vysvětlit diverzifikované strategie v klíčení semen vlastnosti (Simons, 2011), jako je to prokázáno tím, že sbírky se potácel klíčení. Pokud však diverzifikovaná strategie vede jak ke zvýšení průměrné kondice, tak ke snížení rozptylu (např. snížením konkurence mezi sourozenci), pak nepředstavuje zajištění sázek. Podobně, pokud zdánlivá diverzifikace odráží rozdíly mezi jednotlivci, ne uvnitř, pak to není bet-hedging (Starrfelt a Kokko, 2012)., Vyšetřování alpine sazenice rozptýlení vzory a tolerance mrazu a sníh tát, spárované s studium faktorů ovlivňujících klíčení strategie variace v rámci druhu, může pomoci vysvětlit, fenologie klíčení druhů vykazuje rozložené strategie a může indikovat, zda je pravděpodobné, že představují bet-hedging strategií.,
Ekologické Koreláty Klíčení Strategie
Naše zkoumání vztahu mezi klíčení strategie a další ekologicky významné rysy prokázáno, že klíčení rysy byly korelovány s jinými semen rysy, ale ne s vegetativní znaky. Sommerville et al. (2013) zjistil, že australské Alpské byliny s non-endospermickými semeny byly s větší pravděpodobností nespavé při rozptýlení. Naše práce podporuje toto zjištění a dále naznačuje, že menší semena jsou s větší pravděpodobností mezi bezprostředními klíčiteli. Ačkoli Sommerville et al., (2013) neměli předpokládat, proč endospermy může být spojeno s spánku, jsme naznačují, že umístění rezerv v dělohy může brzy zlepšit tempo růstu a usazování v bezprostřední klíčení druhů. Naše výsledky neprokázaly žádný vztah mezi světlem požadavek a klidu, i když lehké požadavky se liší mezi druhy. Tam jsou řadu dalších semen rysy, které pufru riziko vyhynutí v proměnné prostředí, například semen mechanismy a semen dlouhověkosti v půdní semenné bance (viz Tielborger et al., 2012 pro recenzi)., V této fázi nemůžeme říci, zda by tyto rysy vykazovaly korelace se strategií klíčení nebo ne.
dále jsme se zeptali, zda strategie klíčení bude korelována s ekologicky důležitými nereprodukčními rysy, jako je specifická plocha listů nebo výška rostlin (Westoby, 1998; Wright et al., 2004). Zejména jsme předpokládal, že bezprostřední klíčících druhů by mohlo ukázat vlastnost hodnoty svědčí o rychlejší tempo růstu (pomocí SLA jako proxy), jak by mohl být nezbytná pro ustavení v krátkém období mezi rozptýlení a nástup zimy., Takové korelace jsme však nenašli. Zda fyziologické a růstové rysy sazenic v raných stádiích založení mohou vykazovat jiné vzory než dospělé vegetativní rysy, je otázkou hodnou zvážení, ale mimo rozsah současné práce.
Konečně jsme zkoumali, zda druhů z vyšších nadmořských výškách, kde vegetační období jsou obecně kratší, může být více pravděpodobné, že vykazují dormance, ale tam byl žádná souvislost mezi buď výškový rozsah, sbírka nadmořských výškách, nebo průměrná nadmořská výška sběru pro tento druh jako celek a klíčení strategie., Společně tyto výsledky naznačují, že zatímco vývoj strategie klíčení je úzce spojen s ekologicky důležitými rysy osiva, zahrnuje do značné míry nezávislou osu na vegetativních nebo distribučních charakteristikách.
Důsledky pro Alpské Rostlin Společenství v Měnícím se Klimatu
Australské alpské ekosystémy, jako v alpských oblastech po celém světě, jsou pod hrozbou změny klimatu, v kombinaci se změnami v ohni frekvence, využití půdy, příliv invazivních druhů, a dopady zvýšené lidské návštěvy., Oteplování spojené se změnou klimatu dochází rychleji nad treeline než v nižších nadmořských výškách a horských oblastech se očekává, aby i nadále zkušenosti výše průměrné oteplování v budoucnu (Kullman, 2004). Schopnost pokračovat v regeneraci prostřednictvím osiva za nových podmínek bude pravděpodobně hrát významnou roli v reakci alpských rostlin na klima a související změny. Kromě toho pozměněné režimy rušení pravděpodobně vyžadují zvýšenou roli obnovy a rehabilitace v regionu., Poznatky zde uvedené jsou proto rozhodující pro zachování a řízení alpine systémů za změnu; mohou pomoci informovat naše předpovědi o tom, jak studovat druhů může být ovlivněna, tvoří základ množení osiva plány pro tyto druhy, a mohou být použity jako vodítko pro budoucí vyšetřování alpine klíčení semen strategie s identifikovanými zachování a řízení výsledků.,
Změny teploty jsou pravděpodobně ovlivní dormance stav osiva, a to jak pre – a post-disperzní, tedy pozměňování a/nebo narušuje klíčení strategie a má za následek posuny v současné fenologie klíčení (Hoyle et al., 2008b). Pro alpské rostliny, oteplování klimatu může vést k posunu od jara do podzimu vznik, především vlivem změn v semen dormance stav a má za následek závažné důsledky pro druhy, v současné době přizpůsobené vznik na jaře (Mondoni et al., 2012)., Další výzkum na to, jak změny v teplotách, které semena zkušenosti post-rozptýlení, snižuje dobu trvání sněhové pokrývky, a zvýšená frekvence extrémních teplotních událostí bude mít vliv na klíčení jsou zapotřebí předvídat, jak druhy s dormance bude reagovat na změny klimatu. Pro ty, alpské druhy, které klíčí na podzim, před mrazem a sněžení, naše výsledky si položit otázku, jak sazenice těchto druhů, které se bude vyrovnat se s předpovídali snížení srážek a zvýšenou frekvenci-sezónní mráz události.,
a Konečně, naše výsledky si položit důležité otázky týkající se variace v rámci druhu, zejména v souvislosti s předpovídání dopadů změny klimatu nebo výběrem semena, která se využívají pro obnovení. Klíčení strategií může lišit i v rámci druhu v závislosti na počtu obyvatel, dobu květu, semen hmoty, rodičovské investice, klima, nadmořská výška, a/nebo maternální prostředí a tato změna se může odrážet genetické, epigenetické a/nebo environmentální faktory (Cochrane et al., 2015)., Zvýšení nadmořské výšky účty pro změnu v germinability a/nebo dormance stav několika alpských druhů (Billings a Mooney, 1968; Dorne, 1981; Cavieres a Arroyo, 2000), ale vidět (Hoyle et al., 2014). Snížený stav klidu může být důsledkem předčasného ukončení vývoje osiva v důsledku environmentálních stresů (Steadman et al., 2006; Hoyle et al., 2008b)., Vyšetření variability v klíčení strategie v rámci druhu může odhalit potenciál pro druhy, aby vyrovnat se s, a adaptovat se na změny v klimatu, to znamená, že hraje důležitou roli v určování budoucího přežití a rozšíření druhů a poskytuje důležité informace pro řízení a zachování.
Závěry
Náš přístup odhalila rozšířený výskyt dormance v Australské alpské flóry a ukazuje, že klíčení strategie se liší v rámci sběr semen, stejně jako mezi sbírkami., Dále jsme ukázali, že zatímco strategie dormance je fylogeneticky konzervována a korelována s hmotností semen a endospermií, je do značné míry nezávislá na vegetativních vlastnostech a charakteristikách rozsahu. Zatímco z našich výsledků nelze přímo extrapolovat, jak klima a související změny ovlivní alpskou flóru, tyto výsledky mohou tvořit základ pro návrh plánů šíření a další experimentování s dopadem měnících se klimatických režimů., Zejména obhajujeme další výzkum pochopení příčin variací v rámci druhu ve strategii klíčení a role, kterou to může hrát ve schopnosti druhů odolávat předpovídané změně klimatu.
Data o Přístupnosti
Plné klíčení údaje a vlastnost databáze bude podáno na DRYAD a s Atlasu Living Austrálie.
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.,
uznání
uznáváme financování od australské výzkumné rady (LP0991593) a Australské národní botanické zahrady a jejích přátel. Děkujeme J. McAuliffe, s. Fethers a s.Lawatsch z ANBG. Děkujeme také m. Holloway-Phillips, m. Cardillo a s.Cunninghamovi za vstup do dřívějších verzí rukopisu a za analytické rady a s. Venn za přístup k datům rysů.
doplňkový materiál
Foley, M. E. (1992). Účinek rozpustných cukrů a kyseliny giberellové při lámání klidu vyříznutých embryí divokého ovsa (Avena fatua)., Tráva Sci. 40, 208–214.
Google Scholar
Harden, G. J. (1990). Flora Nového Jižního Walesu. Sydney, NSW: Královské botanické zahrady.
Google Scholar
International Seed Testing Association. (2003). Pracovní listy na testování Tetrazolium. Bassersdorf: ISTA.
Kullman, L. (2004). Měnící se tvář alpského světa. Globus. Změna Newsl. 57, 12–14.
Google Scholar
Napsat komentář