
Genetický polymorfismus je existence alternativních státy DNA, určení odchylky organismu je vyšší úroveň integrace. Existují různé druhy modifikací genomu (mutace). Nejvíce studované jsou substituce nukleotidů v kódovacích a regulačních oblastech.
Definice
Genetický polymorfismus je existence, v populaci, z několika alternativních státy DNA, nebo alely, v definované pozici v genomu, nebo locusPosition genu na chromozomu., V populační genetice je soubor homologních genů (homologická třída). Dva geny jsou homologní, pokud odpovídají meióze. Množné číslo: loci.. Tato definice má několik aspektů:
(1) Nejprve musí být vlastnost nesena chromozomy a musí být přenosná.
(2), Pak alely musí být homologousTwo chromozomů nebo dva geny jsou homologní, jestli se shodují a navzájem vylučovat z meiózy. pro jejich postavení v genomu, který vylučuje určité typy variací, které budou diskutovány později.,
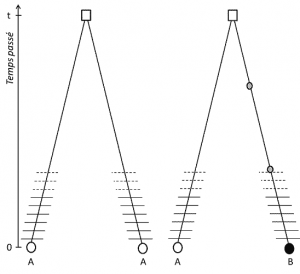
(3) Ale protože rys je přenosné, pozice homologie také znamená, že alely jsou homologní o původu; pokud se liší, je proto, že mutace (alespoň) došlo v jedné z linek, která je spojuje s jejich posledního společného předka (Obrázek 1).
(4) nakonec lze genetický polymorfismus definovat na stupnici nejmenší jednotky tvořící DNA: nukleotidové místo., Jako výsledek, každý nukleotid varianta může určit polymorfismus vyšší úrovně biologické struktury – individuální gen, protein a fenotypu–, které se stanou váhy, na které genetický polymorfismus může být popsán (viz Obrázek 2).
Vezměte si příklad z ABO krevního systému, což odpovídá antigenní motivesPart antigen rozpoznán protilátka nebo lymfocytů receptor. Také známý jako epitop nebo antigenní determinant. Stejný antigen může mít několik epitopů (identických nebo odlišných), a tak vyvolat pestrou imunitní odpověď., podílí se na kompatibilitě během krevních transfuzí. Jedná se o případ genetického polymorfismu přítomného v lidských populacích, zahrnující protein ABO glykosyl transferaseenzym umožňující přenos reziduí obsahujících cukry na proteiny. V případě systému ABO indukuje glykosyltransferáza a A B příslušníka do skupiny a nebo B. pokud jsou přítomny obě glykosyltransferázy, je jedinec skupiny AB. který má tři alely, A, B A O.jedinec může mít (AA), (AO); (BB), (BO); (OO) nebo (AB) genotypy., Jeho fenotyp pak bude ,,; ,;, nebo , kde vidíme, že a a B „ovládnout“ O (tj. jejich vyjádření masky, které o), a že jsou co-dominantní mezi sebou (heterozygotní mezi a a B má rozpoznatelný výraz, ).
polymorfismus lze nejprve popsat na stupnici DNA sekvence lokusu kódujícího protein. Některé polymorfismy jsou „synonymem“, tj. nemění aminokyselinovou sekvenci proteinu; jsou často nejpočetnějšími polymorfismy. Ostatní modifikují aminokyseliny a nazývají se „náhradními“ polymorfismy., Náhradní polymorfismy ABO glykosyl transferázy jsou dva typy: polymorfismy, které mění aminokyselinu, ale ne antigenní jednotky (nejsou v rozporu s ABO fenotyp jedinců); a další náhradní polymorfismy, které určují ABO phenotypeSet pozorovatelných vlastností jedince..
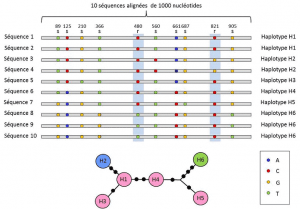
Mezi nukleotidů a alela úrovně genetické analýzy vzít v úvahu střední úroveň popis: lineární uspořádání variabilní místa na místo. Tato uspořádání, nazývaná haplotypyskupina alel různých lokusů umístěných na stejném chromozomu a obvykle přenášených společně. Haplotyp je tvořen kontrakcí anglického výrazu haploidního genotypu nebo haploidního genotypu., Všechny geny umístěné na stejném chromozomu a jejichž alely se oddělují během meiózy, představují haplotyp. Tyto geny se nazývají „geneticky Spojené“, jsou užitečné v evoluční genetice, protože umožňují najít genealogické vazby mezi alely (Obrázek 2).
nukleotidové polymorfismy existují také v intergenních oblastech (mimo kódovací oblasti). Některé ovlivňují oblasti, kde je regulována genová exprese, a proto mají fenotypovou expresi. Jiné nemají žádný známý účinek a jsou označovány jako „tiché“.,
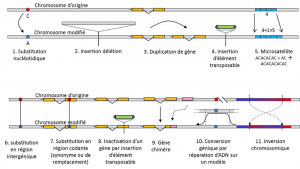
kromě nukleotidové substituce, polymorfismus pro konkrétní typ mutace, mikrosatelitů, odkazuje na rozdíly v délce často nekódující fragment DNA (Obrázek 3) v důsledku změny v počtu opakování v krátké opakované sekvence nukleotidů, např. CACACACA, nebo TGTGTG.
ne všechny genetické variace spadají do definice genetického polymorfismu, protože vyžaduje poziční homologii alternativ (obrázek 3)., Existují tedy opakované sekvence získané duplikací genu, pro který dvě opakování nejsou homologní na pozici. Často nemůžeme mluvit o homologii transpozovatelných prvků, protože obecně mění polohu během buněčných generací, mohou se množit a napadnout genom. Homologii dvou transponovatelných prvků pak nelze definovat. Na druhou stranu, dva kódujících sekvencí stejného lokusu, z nichž jeden je neporušený, a další inaktivován vložení transponovatelných prvků, odpovídají dobře na homologie situace., Jednoho dne, budeme pravděpodobně zvážit mluvit o polymorfismu pro úpravy tzv. „epigenetické“ chromozomů, které jsou někdy předávány přes několik buněčných generací, somaticQualifies non-reprodukční buňky, nebo soma. Somatická mutace, která ovlivňuje Gen somatické buňky, zmizí s nosičem jednotlivce. nebo germinalQualifies gamety. Zárodečná mutace může být přenášena na své potomky., jako je methylace. Jejich význam ve vývoji populací je třeba posoudit., Omezení pojem genetického polymorfismu na určité kategorie variace – v podstatě nukleotidové substituce – pochází z skvělý nástroj posledně v objevování historie přírodních populací. Hodí se k matematickému modelování evoluce.
slovo polymorfismus znamená „několik forem“. Je proti monomorfismu, což naznačuje nepřítomnost variace. Ve slovníku biologie je tato opozice již dlouho používána před genetikou k signalizaci koexistence několika typů jedinců ve stejném druhu, např., kastovní polymorfismus (královna a dělníci) v sociálním hmyzu; sezónní polymorfismus (změny srsti) u některých arktických savců. Tyto případy nespadají pod genetický polymorfismus, novější koncept s užším a přesnějším významem. Kontinuální variace (např. variace velikosti) také nespadají do kategorie polymorfismu, protože nepředstavují odlišné alternativy. Lokus, který ovlivňuje velikost, však spadá do této definice., Níže budeme diskutovat o nejvíce studovaných genetických variacích, protože jsou nejdůležitější pro evoluci, substituce nukleotidů a pak budeme diskutovat o variaci fenotypů.
měření
nestačí říci, že lokus je více či méně variabilní, aniž by tomuto úsudku poskytl kvantitativní odhad. Výzkumník může studovat variaci v různých měřítcích. Pokud se zajímá pouze o alely proteinu, změří alelickou rozmanitost symbolizovanou „H“. Pokud má zájem o DNA rozmanitost, bude měřit nukleotidovou rozmanitost, symbolizovanou „π“.,
alelu rozmanitost H je definována jako pravděpodobnost výkresu dvě různé alely během dvou vzorků s replacementPerforming následné remíze s dodáním p žetonů do urny obsahující n žetony, znamená, že první token, čtení jeho hodnoty, dám ji zpět do urny, přičemž druhý projev, čtení jeho hodnoty, dám ji zpět do urny, atd., až do PTH tokenu. To znamená výběr objektů p mezi n s opakováním (můžete si vybrat stejný objekt několikrát) a v pořadí (pořadí, ve kterém si vyberete objekty, je důležité)., Počet po sobě jdoucích remíz s tokeny mezi n je: n × n × n × … × n = np.. Pokud nazýváme pi frekvencí alely hodnosti i, ukážeme, že pravděpodobnost výběru dvakrát stejné alely je F = Σ pi2. Na allelickou rozmanitost vzorku je pak její doplněk do 1, tedy:
H = 1 – Σ pi2 (1)
To platí jak pro protein alely a haplotypy. Tomu se říká haplotypická rozmanitost. To může být také nazýváno heterozygozitou, protože v případě diploidního lokusu dává očekávanou frekvenci heterozygotů.,
nukleotidová diverzita π je ekvivalentem H na stupnici každého nukleotidu. Je vypočtena jako průměrný počet nukleotidových rozdílů mezi sekvencemi ve vzorku odebraném po dvou (δij), rozdělen podle délky fragment DNA do počtu nukleotidů (L).
π = průměr (δij)/L (2) (viz odkaz )
Tato hodnota se liší mezi druhy. V kódujících částech lidského genomu se dva chromozomy náhodně vybrané z populace liší v průměru o jeden nukleotid na tisíc., V ovocné mušce (Drosophila melanogaster) je tento rozdíl asi jeden ze sta. Moucha je tedy desetkrát variabilnější než člověk. Když tyto hodnoty jsou vztaženy k velikosti kódování regionů (o 15,500 genů Drosophila, alespoň 22,000 u člověka), nebo dokonce genomy (140 milionů párů bází na haploidní genom mouchy, asi 3200 milion nebo dvacet krát více než u člověka), počet polymorfních míst je astronomická a výsledky v tom, že nikdo se rodí z vábivým generace v každém druhu je, byl a bude geneticky totožné s jinou minulost, současnost nebo budoucnost., Informační síla DNA polymorfismu je obrovská. Ve forenzních detektivech mohou detektivové identifikovat každého podezřelého ze šestnácti mikrosatelitů.
historie
termín variace je v názvu prvních dvou kapitol Darwinovy knihy Původ druhů (1859). Darwin zavedl tento pojem do přírodních věd v době, kdy zákony biologické dědičnosti, zůstává záhadou, a věnoval další důležitá kniha, Variace Zvířat a Rostlin (1868)., Přesvědčen o tom, že v evoluci jde o malé změny, které udělal malý rozdíl, adaptace organismů na jejich životní podmínky, byl připojen zásadní význam pro malé kvantitativní změny, které vedly jeho nástupci (zejména Karl Pearson), aby našel biometryScience života měření. Odkazuje ve velmi širokém smyslu na kvantitativní studium živých bytostí.. Znovuobjevení Mendelových zákonů v roce 1900 však přesunulo zájem na diskontinuální variace.
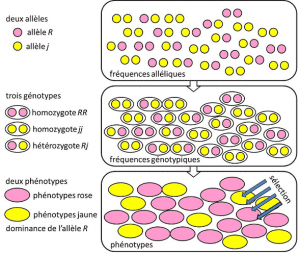
Od roku 1908 do roku 1930, populační genetika byla v podstatě teoretická disciplína se snaží sladit Darwinismus s Mendelovy heredityHeredity založena na přenosu jednoho genu pod dominantní, recesivní nebo sex chromozomu-linked mode X (nebo Y). Odkazuje na genetický rys s jednoduchým determinismem, párem nebo malým počtem párů genů., a kde pravděpodobnosti hrály hlavní roli. Genetika je kontraintuitivní., To předpokládá, že neexistuje žádná reprodukce rodič v jeho potomek, protože ten genotyp výsledků z loterie oddělení alel, než splynutí dvou rodičovské půl genomů. Bylo zjištěno, že se jedná o alelické frekvencefrekvence, při které se alela varianty nachází v populaci. Vyjádřeno jako podíl nebo procento. Součet alelových frekvencí všech alel genu v populaci je tedy podle definice roven 1. V populační genetice představují frekvence alel genetickou rozmanitost na úrovni populace nebo druhu., které jsou přenášeny mezi generacemi, nikoli genotypy nebo fenotypy. Tyto frekvence jsou více či méně stabilní z jedné generace na další a vyrábět, za více zapletení filiations mezi jednotlivci, stejné genotypové frequenciesGenetic struktura obyvatelstva. Určeno z alelických frekvencí (obrázek 4). Distribuce populačních genotypů spojených s rozptylem je proto jediným předvídatelným faktorem., Kolem roku 1930 pomohli tři teoretici Ronald Fisher, JBS Haldane a Sewall Wright vytvořit pojem Mendelovy populacepopulace, jejíž dědičnost poslouchala Mendelovy zákony. . V této souvislosti evoluce nutně spojuje tři strukturální úrovně: Gen, jednotlivce a populaci. Tuto trojcestnou asociaci lze shrnout podle následujícího vzorce: populační evoluce je změna frekvence alel (genová stupnice, ale také skupinová stupnice), změna, která závisí na třídění fenotypů výběrem (individuální stupnice)., Empirické populační genetika výzkum pak vyvinul, ale byl omezen na dlouhou dobu v naší neznalosti, jak chromozomů práce (struktura DNA byla chápána v roce 1953 a její sekvenování začala v roce 1977), a musel ustoupit na studium málo viditelné polymorfismy, jako je zbarvení slunéčko sedmitečné je krovky nebo ulity šneků (viz Zaměření velký šnek rozprava). Mezi genetiky se hodně diskutovalo o tom, zda jsou přírodní populace obecně polymorfní nebo monomorfní, a zda polymorfismus byl sám o sobě prospěšný nebo ne., Studie makromolekulární variace stupnice musela počkat do roku 1966 (pro proteiny) a 1983 (pro DNA), než odpověděla na tyto otázky.
kvantitativní variace a dědičnost
když se rozhlédneme kolem sebe, vidíme fyzické rozdíly mezi bytostmi, některé složité, jako jsou rysy obličeje, jiné snadno měří jako hmotnost nebo výška. Zdravý rozum naznačuje, že jsou částečně přenosné, i když způsobem, který je obtížné specifikovat. Tyto rysy jsou často polygenicképod závislostí mnoha genů. Mluvíme o polygenní dědičnosti. Diabetes je polygenní onemocnění.,, což znamená, že jsou ovlivněny mnoha locuses. Vývojové genetiky, a jeho aplikace na srovnání druhů (evo-devo, pro „vývoj rozvoje“) odhalili síť komplexních interakcí, které dělají, že během vývoje buňky v těle, to vše s identickou genetickou zavazadel, rozlišovat sebe tím, že epigenesis dát různých tkání. Stále není jasné, jak složité postavy jsou postaveny. Měřitelné fenotypy lze studovat pomocí biometrie na základě vlastností statistických distribucí., Jedna hodnota s užitečnými vlastnostmi je rozdílve vzorku jedinců, u kterých byla měřena daná vlastnost, je rozptyl rozdílem mezi průměrem čtverce hodnot a čtvercem průměru hodnot. Toto opatření, které je vždy pozitivní, naznačuje rozptýlení jednotlivců. . Když několik nezávislých příčin určuje změnu čáry, jejich odchylky jsou aditivní a jejich součet dává rozptyl čáry. Pokud nejsou nezávislé, součet kovariancí se přidá k součtu rozptylů., Celkový rozptyl Vt z fenotypová vlastnost je taková, že :
Vt = Vga+Vgd+Vgd+Vgi+Ve
Vga, aditivní genetický rozptyl je součtem rozptylu kvůli každé aktivní nezávisle na ostatních; Vgd, dominantní genetická variance, vyplývá z interakce dvě alely téhož lokusu; Vgi, interakce rozptyl mezi locuses, výsledky ze skutečnosti, že locuses o stejné jedince komunikovat; Ve, rozptyl prostředí, se předpokládá, že zde nezávislé genetické variance.,
genetické variance dominance je společné pro dvě děti stejných rodičů, protože sdílejí dominance účinky, když se v daném lokusu se jim dostalo stejné alely od obou rodičů, který se jim stane na jeden ze čtyř locus. Tyto účinky nesdílejí se svými rodiči, takže jsou si navzájem „podobnější“ než svým rodičům, a to navzdory skutečnosti, že mezi sebou sdílejí tolik genů. Jistě, rodič může náhodou obdržet stejné dvě alely jako jedno ze svých dětí v daném místě., Tato šance závisí na frekvenci alel v populaci. Vidíme, že míra podobnosti mezi dítětem a jeho rodičem pro danou vlastnost je veřejným majetkem.
obecněji se všechny složky genetické rozptylu (Vga+VGD + Vgi) mohou mezi populacemi lišit. V chovných liniích je běžným případem inbreeding, který vyčerpává genetickou odchylku: v důsledku toho může mít vlastnost stejný průměr ve dvou populacích, ale má jinou genetickou variaci. Schopnost chovatele vybrat vlastnost je proto charakteristická pro linii, nikoli vlastnost.,
vztah mezi rodiči a dětmi se měří dědičností h2. Je definován jako poměr aditivní genetické variance na celkové varianci :
h2 = Vga/Vt

(2) korelace 0, kde velikost dětí je nezávislá na velikosti rodičů. Cenný průměrný rodič vytvořil dítě s lhostejnou hodnotou v rámci distribuce dětí. Souřadnice bodů jsou umístěny na kruhu; očekávání jejich hodnoty je umístěno na přímce rovnoběžné s osou rodičů (d)., (3) mezilehlá korelace mezi rodičem a dítětem mezi 0 a 1: souřadnice bodů jsou umístěny na elipse. Očekávání, že jejich hodnota se již nachází na sečna souřadných os (což je také hlavní osa distribuční elipsy), ale na regresní přímce r. Průměrný rodič hodnota produkoval dítě hodnotu b (mezi 0 a) s určitým rozptylem. Je třeba poznamenat, že případ (3) je přechodný mezi případy (1) a (2)
je zobrazen pod různými předpoklady na obrázku 5., Dědičnost je schopnost určit, zda lze funkci Vybrat. Obrázek 5-3 ukazuje, že chovatel bude moci, v křížení generace, přesunout průměrné postavy z hodnoty 0 na hodnotu b, pokud se vybere jedinci s hodnotou. Můžeme ukázat, že b = h2a. Vyberte vlastnost, to je užitečné pro zvýšení dědivosti poměr díky standardizaci prostředí (čímž se sníží environmentální variance), a tím sebe sama v podmínkách prostředí, které upřednostňují vzhled vlastnost., Genetik Waddington ukázal, že v přírodě může změna prostředí odhalit nové rysy, které pak budou vybrány během evoluce.
odkazy a poznámky
obrázek obálky. Šnečí mušle Cepaea nemoralis & Cepaea hortensis. André Künzelmann, UFZ.,
Tento vzorec může být napsán ve více konvenčním způsobem jako: \pi =\frac{n(n-1))}{2}\sum _{i=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
to, Že vzorek jednotlivců, u nichž studoval vlastnost byla měřena, rozptyl lze definovat jako rozdíl mezi tím náměstí hodnot a náměstí střední hodnoty. Toto opatření, které je vždy pozitivní, naznačuje rozptýlení jednotlivců.
to je dědičnost v přísném slova smyslu. Dědičnost v širším smyslu je poměr součtu genetických odchylek k celkové rozptylu., Tento vzorec dává dědičnost u obou rodičů („průměrný rodič“); odhad s jediným rodičem by dal h2/2.
Environmentální Encyklopedie Prostředí Association des Encyclopédies de l ‚Environnement et de l‘ Énergie (www.a3e.fr), smluvně vázáno na University of Grenoble Alpes a Grenoble INP a sponzorované francouzské Akademie Věd.
citovat tento článek:VEUILLE Michel (2021), Genetický polymorfismus a variace, Encyklopedie životního Prostředí, url : https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
články v Encyklopedii životního Prostředí jsou k dispozici pod podmínkami Creative Commons BY-NC-SA licence, která opravňuje reprodukce předmět: citovat zdroje, nikoli komerční použití, sdílení identické počáteční podmínky, reprodukci při každé opětovné použití nebo distribuce zmínka o této Creative Commons BY-NC-SA.
Napsat komentář