DNA og RNA
De to vigtigste typer af nukleinsyrer er deoxyribonukleinsyre (DNA) og ribonukleinsyre (RNA). DNA er det genetiske materiale, der findes i alle levende organismer, lige fra encellede bakterier til multicellulære pattedyr. DNA findes i kernen af eukaryoter og i organeller, chloroplaster og mitokondrier. I prokaryoter er DNA ‘ et ikke lukket i en membranøs konvolut.
hele det genetiske indhold af en celle er kendt som dets genom, og undersøgelsen af genomer er genomik., I eukaryote celler, men ikke i prokaryoter, danner DNA et kompleks med histonproteiner til dannelse af kromatin, substansen af eukaryote kromosomer. Et kromosom kan indeholde titusinder af gener. Mange gener indeholder oplysninger til fremstilling af proteinprodukter; andre gener kode for RNA-produkter. DNA styrer alle de cellulære aktiviteter ved at dreje generne “on” eller “off.”
den anden type nukleinsyre, RNA, er for det meste involveret i proteinsyntese. DNA-molekylerne forlader aldrig kernen, men bruger i stedet en formidler til at kommunikere med resten af cellen., Denne formidler er messenger RNA (mRNA). Andre typer af RNA-lignende rRNA, tRNA og microRNA-er involveret i proteinsyntese og dens regulering.
DNA og RNA består af monomerer kendt som nukleotider. Nukleotiderne kombineres med hinanden for at danne et polynukleotid, DNA eller RNA. Hver nukleotid består af tre komponenter: en kvælstofholdig base, en pentose (fem-carbon) sukker, og en fosfatgruppe (Figur \(\PageIndex{1}\)). Hver nitrogenholdig base i et nukleotid er bundet til et sukkermolekyle, der er bundet til en eller flere fosfatgrupper.,
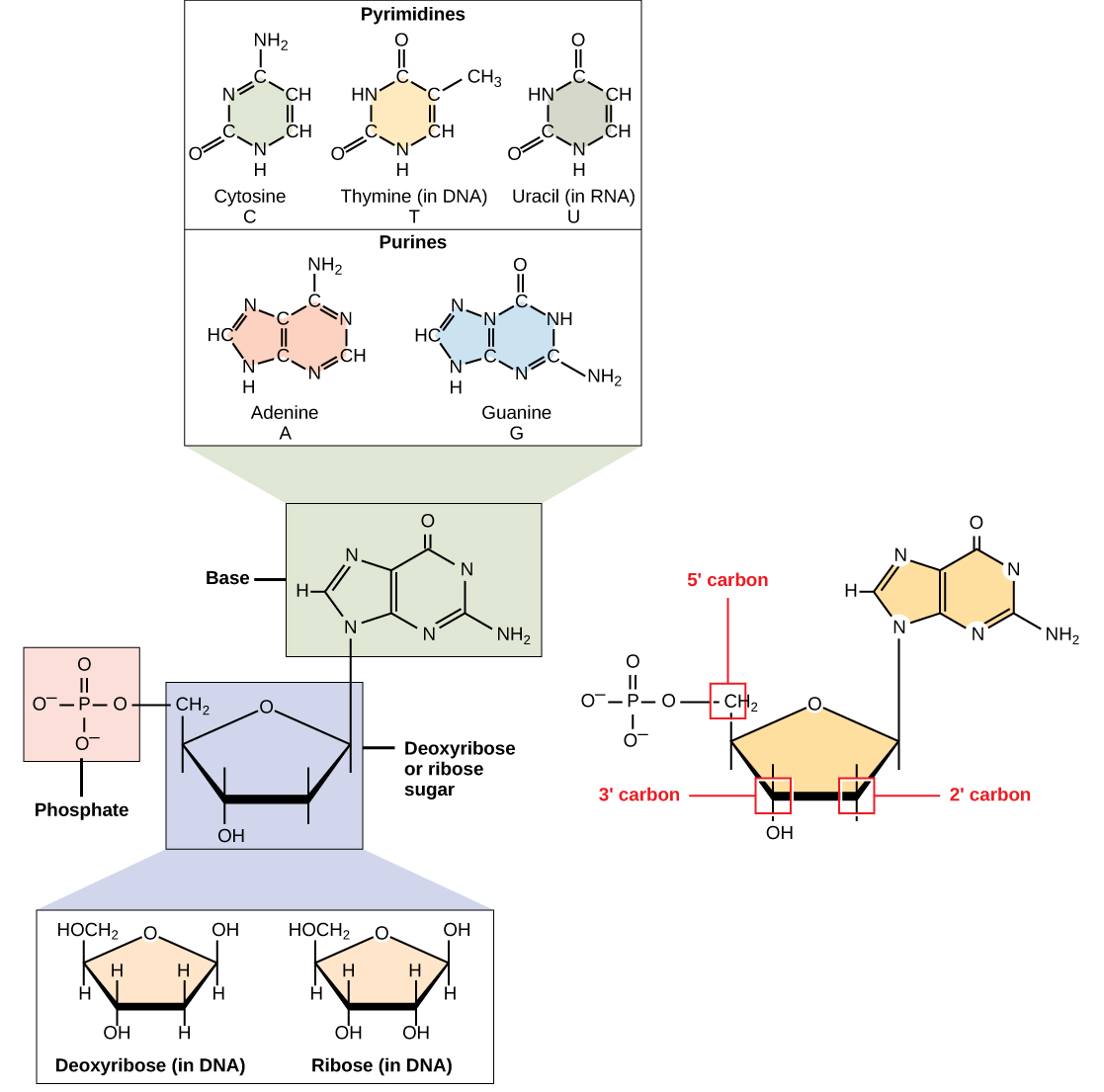
de nitrogenholdige baser, vigtige komponenter i nukleotider, er organiske molekyler og er så navngivet, fordi de indeholder kulstof og nitrogen. De er baser, fordi de indeholder en aminogruppe, der har potentialet til at binde et ekstra brint, og dermed reducerer hydrogenionkoncentrationen i sit miljø, hvilket gør det mere grundlæggende. Hvert nukleotid i DNA indeholder en af fire mulige nitrogenholdige baser: adenin (a), guanin (g) cytosin (C) og thymin (T).
adenin og guanin er klassificeret som puriner., Den primære struktur af en purin er to carbon-nitrogen ringe. Cytosin, thymin og uracil klassificeres som pyrimidiner, der har en enkelt carbon-nitrogenring som deres primære struktur (figur \(\Pageinde. {1}\)). Hver af disse grundlæggende carbon-nitrogen ringe har forskellige funktionelle grupper knyttet til det. I molekylær biologi, stenografi, de kvælstofholdige baser blot kendt ved at deres symboler, A, T, G, C, og U. DNA indeholder A, T, G og C, der henviser til, at RNA indeholder A, U, G og C.
pentose sukker deoxyribose i DNA og RNA, sukker er ribose (Figur \(\PageIndex{1}\))., Forskellen mellem sukkerne er tilstedeværelsen af hydro .ylgruppen på det andet carbon af ribose og hydrogen på det andet carbon af DEO .yribosen (så deo .yribose “mangler” an-OH-gruppen). Carbonatomerne i sukkermolekylet er nummereret som 1′, 2′, 3′, 4′, og 5′ (1 ‘læses som ” en prime”). Phosphat rest er knyttet til hydroxyl gruppe af de 5′ carbon én sukker og hydroxyl gruppe af 3′ carbon sukker af de næste nukleotid, der danner 5′-3’ phosphodiester kobling., Phosphodiesterforbindelsen dannes ikke ved simpel dehydreringsreaktion som de andre forbindelser, der forbinder monomerer i makromolekyler: dens dannelse involverer fjernelse af to fosfatgrupper. Et polynukleotid kan have tusinder af sådanne phosphodiesterforbindelser.
Chargaff ‘ s Regler
Da Watson og Crick, der er fastsat i 1940’erne for at fastlægge strukturen af DNA, var det allerede kendt, at DNA er opbygget af en serie på fire forskellige typer af molekyler, kaldet baser eller nukleotider: adenin (A), cytosin (C), thymin (T), guanin (G)., Watatson og Crick vidste også om Chargaffs regler, som var et sæt observationer om den relative mængde af hvert nukleotid, der var til stede i næsten ethvert ekstrakt af DNA. Chargaff havde observeret, at for en given art, den overflod af var det samme som T, og G var den samme som C. Dette var af afgørende betydning, at Watson & Crick ‘ s model.
Eksempel \(\PageIndex{1}\)
Chargaff bestemmes sammensætningen af nukleinsyrer i prøver fra en række forskellige arter, herunder prokaryotes og eukaryoter. I en bakterieprøve var andelen af adenin 15.,5% (data tilpasset fra Vischer et al, 1949). Hvilken andel guanin ville have været til stede i denne prøve, og hvorfor?
opløsning
fordi et par med T, bør mængden af T være omtrent lig med A, eller cirka 15,5% procent. Således A + T = 15,5 + 15,5 = 31%.
procentdelen af G + C = 100% – 31% = 69%. Fordi G par med C, skal mængden af hver af disse være omtrent ens, så cirka 34, 5% hver.,
DNA Dobbelt Helix Struktur
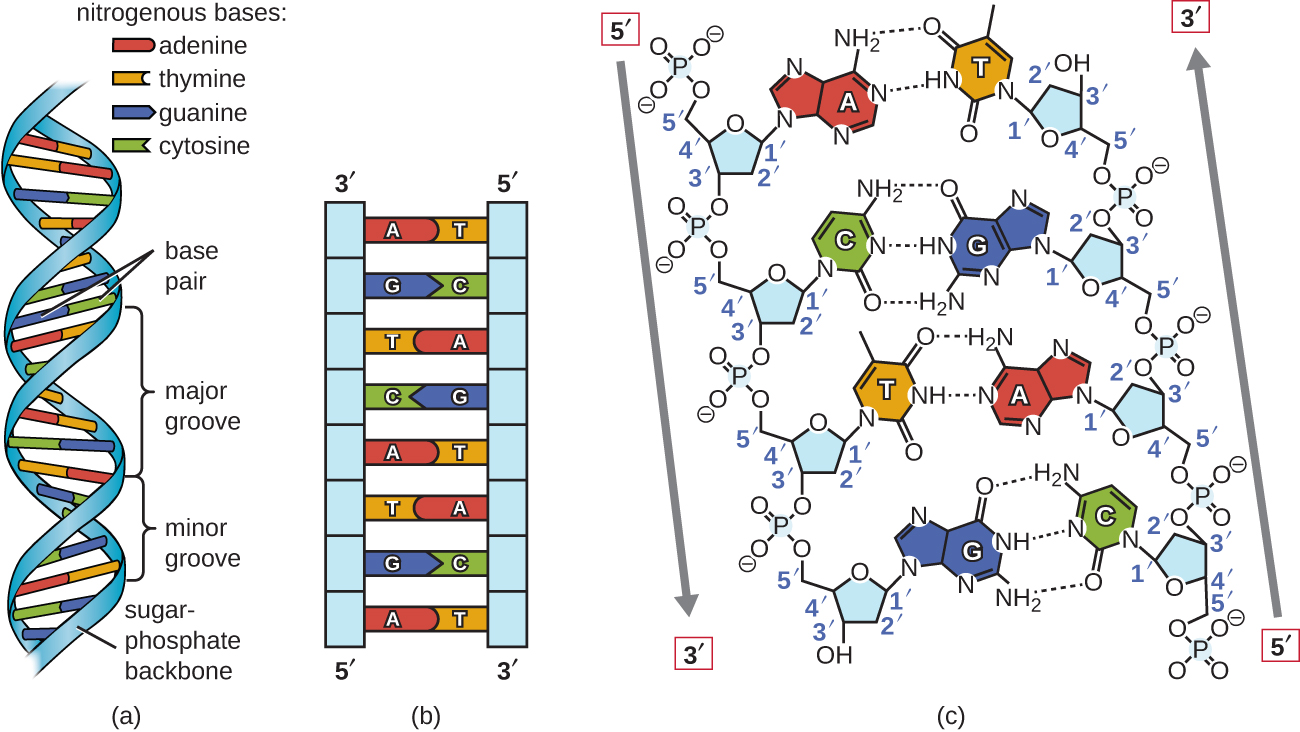
Brug proportional metal modeller af enkelte nukleotider, Watson og Crick udledes en struktur for DNA, der var i overensstemmelse med Chargaff ‘ s Regler, og med x-ray krystallografi data, der blev opnået (med nogle kontroverser) fra en anden forsker ved navn Rosalind Franklin. I Watatson og Cricks berømte dobbelt Heli.indeholder hver af de to tråde DNA-Baser forbundet via kovalente bindinger til en sukkerphosphat-rygrad., Fordi den ene side af hvert sukkermolekyle altid er forbundet med den modsatte side af det næste sukkermolekyle, har hver DNA-streng polaritet: disse kaldes 5′ (5-prime) ende og 3′ (3-prime) ende i overensstemmelse med nomenklaturen for carbonatomer i sukkeret. De to tråde af den dobbelte Heli.løber i anti-parallelle (dvs. modsatte) retninger, med 5′ – enden af en streng ved siden af 3’ – enden af den anden streng. Den dobbelte Heli.har en højrehåndet t .ist (snarere end den venstrehåndede t .ist, der ofte er repræsenteret forkert i populære medier)., DNA-baserne strækker sig fra rygraden mod midten af Heli .en, med et par baser fra hver streng, der danner hydrogenbindinger, der hjælper med at holde de to tråde sammen. Under de fleste forhold er de to tråde lidt forskudt, hvilket skaber en stor rille på den ene side af den dobbelte Heli.og en mindre rille på den anden. På grund af basernes struktur kan A kun danne hydrogenbindinger med T, og G kan kun danne hydrogenbindinger med C (husk Chargaffs regler)., Hver streng siges derfor at være komplementær til den anden, og derfor indeholder hver streng også tilstrækkelig information til at fungere som en skabelon til syntesen af den anden. Denne komplementære redundans er vigtig i DNA-replikation og reparation.
kun visse typer baseparring er tilladt. For eksempel kan en bestemt purin kun parre med en bestemt pyrimidin. Dette betyder, at en kan parre med T, og G kan parre med C, som vist i figur \(\Pageinde. {2}\). Dette er kendt som basen supplerende regel. Med andre ord er DNA-strengene komplementære til hinanden., Hvis sekvensen af en streng er AATTGGCC, ville den komplementære streng have sekvensen TTAACCGG. Under DNA-replikation kopieres hver streng, hvilket resulterer i en datter-DNA-dobbelt Heli.indeholdende en forældres DNA-streng og en nyligt syntetiseret streng.
Tænker fremad motion \(\PageIndex{1}\)
En mutation opstår, og cytosin er erstattet med adenin. Hvilken indflydelse tror du, at dette vil have på DNA-strukturen?,
svar
adenin er større end cytosin og vil ikke være i stand til at basere par korrekt med guaninen på den modsatte streng. Dette vil få DNA ‘ et til at bule ud. DNA reparation en .ymer kan genkende bule og erstatte den forkerte nukleotid.
Skriv et svar