
genetisk polymorfisme er eksistensen af alternative DNA-tilstande, der bestemmer variationen af organismens højere integrationsniveauer. Der er forskellige former for genom modifikationer (mutationer). De mest undersøgte er nukleotidsubstitutioner i kodnings-og reguleringsregioner.
Definition
Genetisk polymorfi er eksistensen, i en befolkning, af flere alternative stater i DNA, eller alleler, i en defineret position i genomet, eller locusPosition af genet på kromosom., I populationsgenetik, et sæt homologe gener (homologiklasse). To gener er homologe, hvis de matcher meiose. Flertal: loci.. Denne definition har flere aspekter:
(1) For det første skal egenskaben bæres af kromosomerne og være overførbar.
(2) så skal allelerne være homologo kromosomer eller to gener siges at være homologe, hvis de matcher og gensidigt udelukker hinanden fra meiose. for deres position i genomet, hvilket udelukker visse typer variation, der vil blive diskuteret senere.,
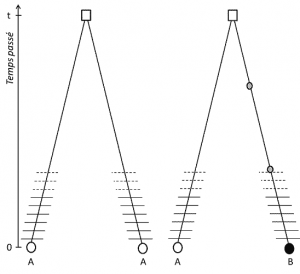
(3) Men da det træk, er overførbare, position homologi indebærer også, at alleler er homologe med afstamning; hvis de er forskellige, det er derfor, at en mutation (mindst) har fundet sted i en af de linjer, der forbinder dem til deres sidste fælles forfader (Figur 1).
(4) Endelig kan genetisk polymorfisme defineres på skalaen af den mindste enhed, der komponerer DNA: nukleotidstedet., Som et resultat kan hver nukleotidvariant bestemme polymorfismen af de højere niveauer af biologisk struktur – individets gen, protein og fænotype – som bliver skalaer, hvor genetisk polymorfisme kan beskrives (figur 2).
Tag eksemplet på ABO-blodsystemet, hvilket svarer til antigene motiveren del af antigenet anerkendt af et antistof eller lymfocytreceptor. Også kendt som en epitop eller antigen determinant. Det samme antigen kan have flere epitoper (identiske eller forskellige) og inducerer således et varieret immunrespons., involveret i kompatibiliteten under blodtransfusioner. Dette er et tilfælde af genetisk polymorfisme, der er til stede i humane populationer, der involverer proteinet ABO glycosyltransferaseen .ym, der tillader overførsel af rester, der indeholder sukkerarter, til proteiner. I tilfælde af ABO-systemet inducerer henholdsvis glycosyltransferase A og B et individ til at tilhøre gruppe A eller B. Når begge glycosyltransferaser er til stede, er individet af gruppe AB. der er tre alleler, A, B og O. En person kan have (AA), (AO); (BB), (BO); (OO) eller (AB) genotyper., Dens fænotype vil så være,,; ,;, eller, hvis vi kan se, at A og B “dominere” O (dvs deres udtryk masker, der O), og at de er co-dominant indbyrdes (den heterozygote mellem A og B, har et genkendeligt udtryk ).
en polymorfisme kan først beskrives ved skalaen af DNA-sekvensen af locus, der koder for et protein. Nogle polymorfismer er” synonyme”, dvs.de ændrer ikke proteinets aminosyresekvens; de er ofte de mest talrige polymorfismer. De andre ændrer aminosyrer og kaldes” erstatning ” polymorfier., Udskiftning polymorfier af ABO glycosyl transferase er af to typer: polymorfier at ændre amino syre, men ikke antigen enhed (de ikke interfererer med den ABO fænotype af personer); og andre udskiftning polymorfier, som bestemmer, ABO phenotypeSet af observerbare karakteristika af en enkeltperson..
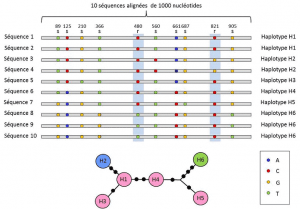
Mellem nukleotid og allel niveauer, genetiske analyser, der tager hensyn til et mellemliggende niveau beskrivelse: den lineære ordning af variable steder på locus. Disse arrangementer, kaldet haplotypergruppe af alleler af forskellige loci placeret på samme kromosom og normalt overføres sammen. Haplotype er en dannet ved sammentrækning af den engelske sætning haploid genotype, eller haploid genotype., Alle gener placeret på det samme kromosom, og hvis alleler adskiller sig sammen under meiose udgør en haplotype. Disse gener kaldes “genetisk forbundet”, er nyttige i evolutionær genetik, fordi de giver mulighed for at finde de genealogiske forbindelser mellem allelerne (figur 2).
nukleotidpolymorfier findes også i intergeniske regioner (uden for kodende regioner). Nogle påvirker de områder, hvor genekspression er reguleret og derfor har fænotypisk ekspression. Andre har ingen kendt virkning og kaldes “tavse”.,
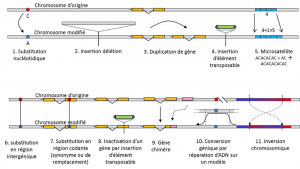
foruden nukleotidsubstitutioner henviser polymorfisme for en bestemt type mutation, mikrosatellitter, til variationer i længden af et ofte ikke-kodende DNA-fragment (figur 3) på grund af en ændring i antallet af gentagelser i en kort gentagen sekvens af nukleotider, f.eks.
ikke al genetisk variation falder inden for definitionen af genetisk polymorfisme, da det kræver positionshomologi af alternativer (figur 3)., Der er således gentagne sekvenser opnået ved duplikering af et gen, for hvilket to gentagelser ikke er homologe pr. Ofte kan vi heller ikke tale om homologi for transposerbare elementer, fordi de generelt ændrer position under cellegenerationer, kan formere sig og invadere genomet. Homologi af to transposable elementer kan ikke derefter defineres. På den anden side svarer to kodningssekvenser af det samme locus, hvoraf den ene er intakt, og den anden inaktiveres ved indsættelse af et transposerbart element, godt til en homologisituation., En dag vil vi sandsynligvis overveje at tale om polymorfisme til modifikationer af såkaldte” epigenetiske ” kromosomer, som undertiden overføres over flere cellegenerationer, somatiskkvalificerer ikke-reproduktive celler eller soma. En somatisk mutation, der påvirker et gen af en somatisk celle, forsvinder med bærerindividet. eller germinalkvalificerer gameter. En kimmutation kan overføres til dets afkom., såsom methylering. Deres betydning i udviklingen af populationer skal stadig vurderes., Begrænsningen af begrebet genetisk polymorfisme til visse kategorier af variation – i det væsentlige nukleotidsubstitutioner – kommer fra sidstnævnte store nytteværdi ved at udforske naturlige populationers historie. De egner sig til matematisk modellering af evolution.
ordet polymorfisme betyder “flere former”. Det er imod monomorfisme, hvilket indikerer fraværet af variation. I biologiens ordforråd er denne opposition længe blevet brugt før genetik til at signalere sameksistensen af flere typer individer i samme art, f. eks., kastepolymorfisme (dronning og arbejdere) i sociale insekter; sæsonbestemt polymorfisme (pelsændringer) hos nogle arktiske pattedyr. Disse tilfælde falder ikke under genetisk polymorfisme, et nyere koncept med en smalere og mere præcis betydning. Kontinuerlige variationer (f.eks.størrelsesvariation) falder heller ikke ind under kategorien polymorfisme, da de ikke præsenterer forskellige alternativer. Locus, der påvirker størrelse, falder dog inden for denne definition., Vi vil diskutere nedenfor de mest studerede genetiske variationer, fordi de er de vigtigste for evolution, nukleotidsubstitutioner, og så vil vi diskutere variationen af fænotyper.
måling
det er ikke nok at sige, at et locus er mere eller mindre variabelt uden at give et kvantitativt skøn til denne dom. En forsker kan studere variationen i forskellige skalaer. Hvis han kun er interesseret i et proteins alleler, måler han den alleliske mangfoldighed, symboliseret med ‘H’. Hvis han er interesseret i DNA-mangfoldighed, vil han måle nukleotiddiversitet, symboliseret med ‘π’.,
Den allel, mangfoldighed H er defineret som sandsynligheden for at trække to forskellige alleler i løbet af to stikprøver med replacementPerforming en efterfølgende tegne med levering af s indsats i en urne, der indeholder n kuponer, betyder at tage et første token, læse dens værdi, sætte det tilbage i en urne, der tager et andet token, læse dens værdi, sætte det tilbage i urnen, osv., indtil pth token. Dette betyder at vælge p-objekter blandt n med gentagelse (du kan vælge det samme objekt flere gange) og i rækkefølge (den rækkefølge, du vælger objekterne, er vigtig)., Antallet af successive træk med tokens blandt n er: n.n. n … … … n = NP.. Hvis vi kalder pi hyppigheden af allelen af rang i, viser vi, at sandsynligheden for at plukke to gange den samme allel er f = pi pi2. Allelerne mangfoldighed af prøven er derefter dens komplement til 1, dvs:
H = 1 – Σ pi2 (1)
Dette gælder for både protein alleler og haplotypes. Dette kaldes haplotypisk mangfoldighed. Det kan også kaldes Hetero .ygositet, fordi det i tilfælde af et diploid locus giver den forventede frekvens af Hetero .ygoter.,
nukleotiddiversiteten is svarer til H ved skalaen af hvert nukleotid. Det beregnes som gennemsnittet af antallet af nukleotidforskelle mellem sekvenser i en prøve taget to og to (δij), divideret med længden af DNA-fragmentet i antallet af nukleotider (L).
π = gennemsnit (ijij) /L (2) (Se reference)
denne værdi adskiller sig mellem arter. I de kodende dele af det humane genom varierer to kromosomer tilfældigt udvalgt fra befolkningen i gennemsnit med et nukleotid pr., I frugtfluen (Drosophila melanogaster) er denne forskel omkring et ud af hundrede. Fluen er således ti gange mere variabel end mennesket. 15.500 gener i Drosophila, mindst 22.000 hos mennesker) eller endda genomer (140 millioner basepar pr.haploid genom i fluer, omkring 3.200 millioner eller tyve gange mere hos mennesker), er antallet af polymorfe steder astronomisk og resulterer i, at ingen bliver født af den seksualiserede generation i nogen art er, har været og vil være genetisk identisk med en anden fortid, nutid eller fremtid., Informationskraften af DNA-polymorfisme er enorm. I kriminalteknik kan detektiver identificere enhver mistænkt fra seksten mikrosatellit-lokuser.
historie
udtrykket variation er i titlen på de to første kapitler i Dar .ins bog The Origin of Species (1859). Dar .in introducerede denne opfattelse i naturvidenskaben på et tidspunkt, hvor lovene om biologisk arvelighed forblev et mysterium og viet en anden vigtig bog til det, Variation in Animals and Plants (1868)., Overbevist om, at evolutionen handlede om små variationer, der gjorde ringe forskel for tilpasningen af organismer til deres levevilkår, lagde han afgørende vægt på små kvantitative variationer, hvilket fik hans efterfølgere (især Karl Pearson) til at finde biometryScience of life measurement. Henviser i meget bred forstand til den kvantitative undersøgelse af levende væsener.. Men genopdagelsen af Mendels love i 1900 skiftede interesse for diskontinuerlige variationer.
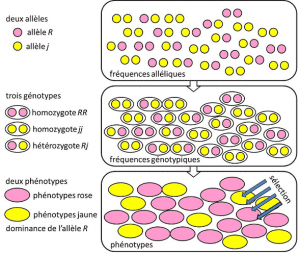
Fra 1908 til 1930, populationsgenetik var hovedsageligt en teoretisk disciplin, der søger at forene Darwinismen med Mendelian heredityHeredity baseret på transmission af et enkelt gen, under en dominant, recessiv eller kønskromosom-forbundet mode X (eller Y). Henviser til et genetisk træk med simpel determinisme, af et par eller et lille antal par gener., og hvor sandsynligheder spillede en stor rolle. Genetik er counter-intuitiv., Det forudsiger, at der ikke er nogen reproduktion af forældrene i hans efterkommer, fordi sidstnævntes genotype er resultatet af lotteriet om adskillelse af allelerne før fusionen af de to forældres halvgenomer. Det blev indset, at disse er de alleliske hyppighederfrekvens, hvor allelen af en variant findes i en befolkning. Udtrykt som en andel eller procentdel. Summen af allelfrekvenserne for alle alleler af et gen i en population er derfor per definition lig med 1. I populationsgenetik repræsenterer allelfrekvenser genetisk mangfoldighed på populations-eller artsniveau., der overføres mellem generationer, ikke genotyper eller fænotyper. Disse frekvenser er mere eller mindre stabile fra en generation til den næste og producerer, ud over den multiple sammenfiltring af filiationer mellem individer, den samme genotypiske frekvensgenetiske struktur af befolkningen. Bestemt ud fra de alleliske frekvenser (figur 4). Fordelingen af populationsgenotyper, der er forbundet med varians, er derfor den eneste forudsigelige faktor., Omkring 1930, de tre teoretikere Ronald Fisher, JBS Haldane og Sewall Wright var med til at skabe forestillingen om Mendelian populationPopulation hvis arvelighed adlød mendels love. . I denne sammenhæng forbinder evolution nødvendigvis tre strukturelle niveauer: genet, individet og befolkningen. Denne trevejsforening kan opsummeres med følgende formel: befolkningsudvikling er en ændring i allelfrekvens (genskala, men også gruppeskala), en ændring, der afhænger af sorteringen af fænotyper ved udvælgelse (individuel skala)., Empiriske populationsgenetik forskning, så udviklede, men var begrænset til en lang tid af vores uvidenhed om, hvordan kromosomer arbejde (DNA-strukturen blev forstået i 1953 og dens sekventering begyndte i 1977), og var nødt til at falde tilbage på studiet af de få synlige polymorfier, såsom farvning af mariehøne er elytra eller skallen af snegle (se Fokus Den store snegl forhandling). Der var meget debat blandt genetikere om, hvorvidt naturlige populationer generelt var polymorfe eller monomorfe, og om polymorfisme i sig selv var gavnlig eller ej., Undersøgelsen af makromolekylær skalavariation måtte vente til 1966 (for proteiner) og 1983 (for DNA), før de besvarede disse spørgsmål.
kvantitative variationer og arvelighed
Når vi ser os omkring, ser vi fysiske forskelle mellem væsener, nogle komplekse som ansigtstræk, andre lette at måle som vægt eller højde. Sund fornuft antyder, at de delvis kan overføres, skønt på en måde, der er vanskelig at specificere. Disse træk er ofte polygeniskeunder afhængighed af mange gener. Vi taler om polygenisk arvelighed. Diabetes er en polygenisk sygdom.,, hvilket betyder, at de er påvirket af mange lokaliteter. Udviklingsmæssige genetik, og dens anvendelse til sammenligning af arter (evo-devo, for “udviklingen i udvikling”) optrævle web af komplekse interaktioner, der gør, at der under udviklingen af de celler i kroppen, alle med samme genetiske bagage, differentiere sig ved at epigenesis til at give forskellige væv. Det er stadig uklart, hvordan komplekse tegn er bygget. Målbare fænotyper kan studeres ved hjælp af biometri, baseret på egenskaberne ved statistiske distributioner., En værdi med nyttige egenskaber er varianseni en prøve af individer, hvor et givet træk blev målt, er variansen forskellen mellem gennemsnittet af kvadratet af værdier og kvadratet af gennemsnittet af værdierne. Denne foranstaltning, som altid er positiv, indikerer spredning af individer. . Når flere uafhængige årsager bestemme variationen af en linje, deres afvigelser er additive og deres sum giver variansen af linjen. Hvis de ikke er uafhængige, tilføjes summen af kovariancerne til summen af afvigelserne., Den samlede varians Vt af en fænotypiske træk er således, at :
Vt = Vga+Vgd+Vgd+Vgi+Ve
Vga, additive genetiske varians, er summen af varianserne på grund af hvert locus uafhængigt af de andre; Vgd, dominerende genetiske varians, er et resultat af samspillet af de to alleler af de samme locus; Vgi, interaktion variansen mellem locuses, resultater fra det faktum, at locuses af samme individ interagerer; Ve, variansen af miljøet, antages her, uafhængig af genetiske varians.,
den genetiske varians af dominans er fælles for to børn af de samme forældre, fordi de deler dominanseffekter, når de på et givet sted har modtaget de samme alleler fra begge forældre, hvilket sker med dem på et ud af fire locus. De deler ikke disse effekter med deres forældre, så de er mere “lignende” til hinanden end de er til deres forældre, på trods af at de deler så mange gener med hinanden. Sikkert, en forælder kan, ved en tilfældighed, har modtaget de samme to alleler som et af hans børn på et givet sted., Denne chance afhænger af hyppigheden af alleler i befolkningen. Vi ser, at graden af lighed mellem et barn og hans eller hendes forælder for en given egenskab er en offentlig ejendom.
mere generelt kan alle komponenter af genetisk varians (VGA+Vgd+Vgi) variere mellem populationer. I avlslinjer er et almindeligt tilfælde indavl, der udtømmer genetisk variation: som et resultat kan et træk have det samme gennemsnit i to populationer, men har en anden genetisk variation. En opdrætteres evne til at vælge et træk er derfor et kendetegn for linjen, ikke for egenskaben.,
forholdet mellem forældre og børn måles ved arvelighed h2. Det er defineret som forholdet mellem additive genetiske varians til den samlede varians :
h2 = Vga/Vt

(2) korrelation af 0, hvor størrelsen af børn er uafhængig af forældrenes. En værdifuld gennemsnitlig forælder har produceret et barn af ligegyldig værdi inden for fordelingen af børn. Koordinaterne for punkterne er placeret på en cirkel; forventningen om deres værdi er placeret på en linje parallelt med forældrenes akse (d)., (3) mellemliggende forældre-barn korrelation mellem 0 og 1: koordinaterne for punkterne er placeret på en ellipse. Forventningen om, at deres værdi er ikke længere placeret på vinkelhalvering af koordinatsystemet akser (der er også den største akse for det distribution ellipse), men på den regressionslinje r. En gennemsnitlig forælder af værdi har produceret et barn af værdi b (mellem 0 og a) med en vis varians. Det bemærkes, at (3) er mellemliggende mellem tilfældene (1) og (2)
Det er vist under forskellige antagelser i Figur 5., Arvelighed er evnen til at afgøre, om en funktion kan vælges. Figur 5-3 viser, at en opdrætter vil være i stand til, inden for en passage generation, til at flytte gennemsnittet af en karakter fra værdien 0 til værdien b, hvis han vælger personer med en værdi. Vi kan vise, at b = h2a. For at vælge en egenskab, er det nyttigt at øge arveligheden forholdet ved at standardisere miljøet (og dermed reducere de miljømæssige varians), og ved at placere sig selv i de miljømæssige forhold, der fremmer forekomsten af de træk., Genetiker .addington har vist, at en miljøændring i naturen således kan afsløre nye træk, der derefter vælges under evolutionen.
referencer og noter
forsidebillede. Sneglehuse Cepaea nemoralis & Cepaea hortensis. Andr.K .n .elmann, UF..,
Denne formel kan være skrevet i en mere konventionel måde: \pi =\frac{n(n-1))}{2L}\sum _{i=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
at Være en stikprøve af personer, hos hvem de undersøgte træk blev målt, variansen kan være defineret som forskellen mellem middelværdien af kvadratet på værdier og kvadratet af gennemsnittet af de værdier. Denne foranstaltning, som altid er positiv, indikerer spredning af individer.
Dette er arvelighed i streng forstand. Arvelighed i bred forstand er forholdet mellem summen af genetiske afvigelser og total varians., Denne formel giver arvelighed med begge forældre (den “gennemsnitlige forælder”); estimatet med kun en forælder ville give h2/2.
De Miljømæssige Encyklopædi af Miljøet ved Association des Encyclopédies de l ‘environnement et de l’ Énergie (www.a3e.fr), kontraktligt knyttet til Universitetet i Grenoble Alpes og Grenoble INP, og sponsoreret af den franske videnskabsakademi.
for at citere denne artikel: VEUILLE Michel (2021), genetisk polymorfisme og variation, Encyclopedia of the Environment, url : https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
artiklerne i Encyclopedia of the Environment stilles til rådighed under betingelserne i Creative Commons BY-NC-SA-licensen, som tillader reproduktion underlagt: citerer kilden, ikke gør kommerciel brug af dem, deler identiske oprindelige betingelser, gengiver ved hver genbrug eller distribution omtalen af denne Creative Commons BY-NC-SA-licens.
Skriv et svar