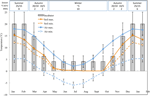
Figur 1. Månedlige maksimale og minimale luft-og jordtemperaturdata i de australske Alper (gennemsnit ± STDEV). Lufttemperaturer er gennemsnit fra Bom-databasen, jordtemperaturer blev målt med iButtons (se metoder). Søjler angiver eksperimentelle inkubator temperaturer (gennemsnit ± STDEV)., Den ækvivalente årstid( sæson), inkubator temperatur regime (12 C, 12/12 h, lys/mørk) og behandlingsvarighed (uger) som angivet i toppanelet.
Med henblik på at afdække temperaturforhold tæt på dem, der frø erfaring post-spredning, daglig maksimum og minimum jordens temperatur data blev indsamlet inden for undersøgelsesområdet (Seaman ‘ s Hytte; 36.27°S og 148.17°E, 2030 m en.s.l.) hjælp ibutton dataloggere (n = 10, Indlejrede datasystemer, USA), placeret 4 cm under jordoverfladen i bunden af vegetation fra 17. januar til 17. December 2012., Vi placerede loggere på 4 cm for at undgå overfladeforstyrrelser, men er stadig repræsentative for forhold, som frø udsættes for i jord; denne dybde er mellemliggende til det, der er blevet brugt i tidligere alpine jordtemperaturovervågningsøvelser (Scherrer and K .rner, 2011; Pauli et al., 2015). Som Mistænkt faldt den gennemsnitlige jordtemperatur ikke under frysepunktet om vinteren (Figur 1). Om sommeren er jordtemperaturen under vegetation kendt for at spore Omgivelsestemperatur, hvorimod temperaturer på bar jord vil overstige omgivelsestemperaturen med op til 15 C C (Soil Conservation Service upublicerede poster, 1960-1970 ‘ erne).,
Indsamling af Frø og Spiring
Modne frø af 54 arter fra 16 familier og 37 slægter blev indsamlet i perioden mellem januar og April 2009, 2010 og 2011 (se Tabel 1 for fuld navne og myndigheder. Værdikuponer blev indgivet til Australian National Herbarium, Canberra). I alt repræsenterede arten mere end en fjerdedel af den australske angiosperm flora, der findes i alpine regioner (Costin et al., 2000, Se tabel 1), skønt mange af disse arter også strækker sig til under treeline., Levedygtigheden af alle samlinger blev estimeret før såning under eksperimentelle spiringsbetingelser ved anvendelse af Tetra .oliumchlorid (t.) farvningsteknik (International Seed Testing Association, 2003). For yderligere oplysninger om indsamling og behandling se tillæg A i supplerende materiale.
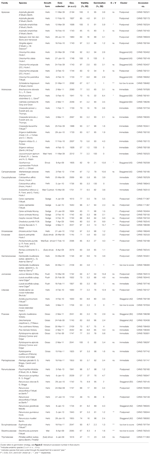
Tabel 1. Undersøgelse arter, familie og myndighed, vækstform og samlinger detaljer, procentdel frø levedygtighed (t.e.-estimeret ,.e. S. E.), endelig procentdel spiring opnået (S S. E.,), statistisk forskel mellem levedygtighed og spiring (g < V, kun testet i tilfælde, hvor spiring var mindre end levedygtighed).
fænologi af frøspiring blev undersøgt ved at efterligne temperaturregimer, som alpine frø oplever in situ, efter spredning, i en kunstigt forkortet progression af årstider. Frø blev indlejret i hele eksperimentet, således at resultaterne indikerer potentiale for spiring, når vand ikke begrænser., Luft-og jordtemperaturdata (Figur 1) blev brugt til at styre og validere inkubatortemperaturregimer, skønt de logistiske kontraindikationer for at arbejde med inkubatorer udelukkede inkorporering af de udsving, der er forbundet med naturlige forhold. Spiring test anvendte otte replikater af 25 frø pr samling, sået i 9 cm diameter plast petriskåle indeholdende 1% almindeligt vand-agar. Petriskåle blev forseglet ved hjælp af Parafilm for at undgå agarudtørring, før de blev anbragt i spiringsinkubatorer (Thermoline Scientific, Melbourne, NS., Australien)., Halvdelen af retterne blev indpakket i aluminiumsfolie for at udelukke lys og give en indikation af lyskrav til spiring. Hver af de fire replikater pr lysbehandling pr indsamling blev anbragt på en anden hylde inden for samme inkubator og placering inden for denne hylde blev re-arrangeret ugentligt. Alle replikater af hver samling blev flyttet gennem følgende simuleret thermoperiods: 8 uger på 25/15°C (dag/nat-temperatur) → 2 uger på 20/10°C → 2 uger på 10/5°C → 10 uger på en konstant 4-5°C → 2 uger på 10/5°C → 2 uger på 20/10°C → 6 uger på 25/15°C., Således efterlignede en cyklus af hele eksperimentet temperaturer, der minder om sommer, tidligt efterår, sent efterår, vinter, tidligt forår, sent forår og sommer og varede i alt 32 uger. Det forkortede år blev betragtet som rimeligt, fordi tiltrædelser generelt var ophørt med yderligere spiring i en given behandling, inden de blev flyttet til den næste “sæson”., En 12/12 h lys/mørke lysperiode blev leveret i løbet af lysstofrør (> 90 μ mol m−2 s−1 i midten af hver hylde), og i løbet af foråret, sommeren og efteråret temperatur regimer lys faldt sammen med den varme temperatur periode af dagen. Spiring, defineret som radicle fremkomst med mere end 1 mm, blev scoret hver 7.dag, og spirede frø og frøplanter blev efterfølgende fjernet fra opvasken. Spiring af folieindpakkede plader blev vurderet som ovenfor, men under meget lavt grønt lys.,efter en forsøgscyklus (32 uger) blev de fleste samlinger afsluttet (se supplerende info for undtagelser), efter at have udvist nul spiring i mindst 2 uger på dette tidspunkt. Efter afslutningen blev alle resterende intakte frø dissekeret med en skalpel under et mikroskop. Frø med en fast, frisk endosperm og embryo blev betragtet som levedygtige, og frø, der var tomme for et embryo, blev fratrukket det samlede beløb ved beregning af procentvis spiring for hver samling., Vi beregnede et indeks over lyskravene til spiring for hver art som en brøkdel af spiring i den mørke behandling på det tidspunkt, hvor kumulative spiring percentiler på 50 og 95% blev nået i den skiftende lys / mørke behandling (se nedenfor).
økologiske korrelater af Spiringsstrategi
vi udarbejdede en database over arternes funktionelle træk og geografiske mål fra offentliggjorte data for at vurdere korrelationer med spiringsstrategi., Databasen omfattede følgende kontinuerlige træk: gennemsnitlig frømasse, lysbehov for spiring (se ovenfor), specifikt bladareal (frisk areal/tørmasse), individuelt bladareal, individuel bladtørvægt og højde ved modenhed. Minimum, gennemsnitlige og maksimale indsamlingsforhøjelser for arten blev bestemt ud fra alle herbariumrekorder i det australske virtuelle Herbarium, og den højde, hvorpå de specifikke frøsamlinger, der blev brugt i undersøgelsen, blev foretaget, blev også inkluderet som kontinuerlige træk. Vækstform blev scoret som en af tre kategorier: graminoid, urt eller busk., Vi inkluderede også et binært træk: om frøene var endospermiske eller ej.
De træk blev indsamlet data fra en række forskellige kilder, herunder Australske Virtuelle Herbarium (http://avh.chah.org.au/), den Australske Nationale Herbarium prøve oplysninger register (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Alpine Planter (Costin et al., 2000), ne.South Floraales Flora (Harden, 1990) og den komparative interne morfologi af frø (Martin, 1946). Frømasse og specifikt bladareal blev opnået fra individuelle feltsamlinger anvendt i spiringsforsøgene., Frømasse blev opnået ved at veje fire partier af 10 frø. SLA blev bestemt baseret på fem fuldt udvidede blade fra separate individer pr. Disse blev indsamlet i området, scannet på en flatbed scanner, tørret ved 60°C til konstant vægt og vejede til den nærmeste 0.000 g. Hvor der er mangel på frø, der var til rådighed, eller specifikke blad område, der ikke var tilgængelige data blev indhentet fra andre ANBG samlinger af samme art eller fra andre datakilder (S. Venn, upublicerede data)., Hvorvidt et frø var endospermisk eller ej, blev bestemt ud fra tegninger af frøene i ANGB seed bank records i samråd med anbg seed bank personale. Den fulde database og kilder er tilgængelig på anmodning fra forfatterne.
Analyse af Data
Spiring mønstre blev vurderet visuelt og ved hjælp af en klyngeanalyse i PC-ORD til Windows, version 6 (BMS-Software Design, Gleneden Beach, ELLER, USA). Analysen brugte en euklidisk afstandsmatri.og en nærmeste nabo sammenføjningsalgoritme., Datapunkter for klyngeanalysen var tiden (ugerne) til specifikke kumulative spiringsprocentiler (T20, T40, T50, t60, t80 og t95%) for hver samling (i gennemsnit på tværs af de 4 retter). Således ville en samling, der spirede tidligt i eksperimentet, have korte gange til percentiler op til t80% for eksempel, hvorimod en samling, der udsatte spiring, indtil temperaturerne steg efter den simulerede vinter, ville have lange gange til disse percentiler., Mere komplekse spiringsmønstre, såsom langsom, stabil spiring eller udbrud af spiring, kunne også påvises, og ville blive indikeret med jævnt fordelte tider eller tider grupperet i henholdsvis starten og slutningen af eksperimentet. Denne metode blev anvendt som en måde at pålægge en objektiv vurdering af spiringsmønster, uafhængigt af temperaturen i sig selv.,
en envejs variansanalyse (ANOVA) blev udført for replikatpladerne i hver samling for at vurdere, om der var en signifikant forskel mellem procentvis levedygtighed (som bestemt ved T.-testen) og endelig procentvis spiring. Yderligere generelle lineære modeller blev brugt til at vurdere, om klyngerne som følge af klyngeanalysen var forskellige i levedygtighed eller endelig spiringsprocent.
foreninger mellem økologiske træk og spiringsstrategieklynger blev testet på to måder., For det første testede vi for gennemsnitlige forskelle mellem klynger ved hjælp af ANOVA med klynge som en fast effekt og økologiske korrelater som responsvariabler eller ved hjælp af replikerede G-test for endospermi og vækstform. For det andet, fordi spiring og andre funktionelle træk meget sandsynligt vil blive påvirket af evolutionær historie, vurderede vi disse korrelationer ved hjælp af fylogenetiske regressioner. En fylogeni blev konstrueret til undersøgelsesarten ved hjælp af Phylocom Phylomatic og bladj-pakningen i R (.ebb et al., 2008; R Kernehold, 2013)., Vi vurderede tilstedeværelsen af fylogenetisk struktur i variabler ved anvendelse af phylosig eller phylo.d (diskrete eller binære variabler) funktion af phytools pakken i R (Revell, 2012). Fylogenetiske regressioner blev udført under anvendelse af pgls-funktionen i Caper (Orme et al., 2013). Analyser blev udført på en struktureret måde, der afspejler den klyngedannelse, der blev afsløret i PC-Ord (se nedenfor)., Oprindeligt inkluderede vi fire spiringsstrategier (udskudt, øjeblikkelig og forskudt kategorier inden for hver af de foregående), derefter tre (klumpede alle forskudte arter og sammenlignede med udskudt og mellemliggende) og endelig vurderede vi forskelle mellem de to bredeste strategier, udskudt og mellemliggende.
resultater
frøets levedygtighed sammenlignet med den endelige spiring
samlet set var frøets levedygtighed og den endelige spiringsprocent høj. Gennemsnitlig t. – estimeret levedygtighed for alle samlinger var 80 1 1.,7%, hvor mere end to tredjedele af samlingerne udviser mere end 75% levedygtighed (tabel 1). Der var ingen forskel i kvaliteten af samlinger på tværs af de 3 år (gennemsnit levedygtighed i 2009, 2010 og 2011 var 79 ± 2.8, 80 ± 2.7, og 81 ± 3.7%, henholdsvis), hvilket indikerer, at den bank af frø indsamlet i 2009 ikke mindske deres levedygtighed.
Spiring, der oversteg eller var ikke væsentligt lavere end TZ-anslået levedygtighed blev opnået for 45 af de 54 arter (One-Way ANOVA: P>0.05, Tabel 1)., Af de resterende ni arter var den endelige spiring af fire arter meget lav (< 5%) på trods af god levedygtighed (ca. 55-94%), og derfor kunne vi ikke drage nogen konklusioner om disse arters spiringsstrategier. Manglende spiring kunne afspejle enten at vores behandlinger ikke lindre dvale eller at frøene ikke var fuldt modne ved indsamling. Skæreprøver ved afslutningen af eksperimentet viste, at størstedelen af ikke-spirede frø i alle tilfælde indeholdt et sundt, imbibed embryo., Interessant nok opnåede 34 arter spiring, der var større end den T. – estimerede levedygtighed (tabel 1), hvilket antyder, at T. – testen ofte undervurderede frøets levedygtighed.
spirende klynger
klyngeanalyse afslørede to store klynger (A og B) med understruktur i hver (figur 2). For størstedelen af arterne (n = 21 arter fra 10 familier) blev spiring udsat indtil i en periode med vintertemperaturer (5 C C Dag/nat) eller indtil temperaturerne blev hævet til 10/5.C efter vinteren (A1 og A1:vinter, figur 2, 3A)., Alternativt begyndte spiring straks ved 25/15.c (n = 17 arter, seks familier, set i klynge B1, figur 2, 3B). For størstedelen af disse arter var spiringshastigheden hurtig; 50% af den samlede spiring (t50) forekom på mindre end 3 uger (supplerende Figur 1).
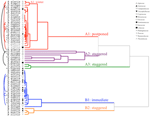
figur 2. Klyngeanalyse dendrogram baseret på tid til kumulative spiring percentiler (20, 40, 50, 60, 80, og 95%). Arter angivet med slægten initial og første del af artsnavnet. Stjerner markerer samlinger, der blev banket før testning., Fine buer forbinder duplikatsamlinger for at sammenligne effekten af bank på spiring. Symboler angiver forskellige familier (se legenden). Samlinger vist i grå tekst er dem, der blev udsat for to cyklusser af eksperimentet.
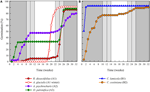
Figur 3. Kumulativ procentvis spiring af en repræsentativ art fra hver spireklynge. (A) klynge a, arter, der udsatte spiring, (B) klynge B, arter, der spirede øjeblikkeligt., Bemærk, at forskudte spiringsmønstre er indlejret i ovennævnte klynger. Inkubator temperaturregimer er repræsenteret ved skygge (se figur 1).
Inden for hver af disse overordnede klasser, men der var klart sub-klynger, der afspejler spiring, der blev forskudt over tid vil sige, at en del af de frø masse spiret før eksponering til vinter temperaturer, mens den resterende del spiret efter vinteren. Forskudt spiring forekom hos 13 arter fra seks familier og forekom primært, når dagtemperaturerne oversteg 10 C C (figur 3A,B)., Disse forskudte spirer blev opdelt i tre grupper i klyngeanalysen. Klynge A2 med syv samlinger viste betydelig spiring både før og efter den kolde temperaturperiode. Hvor spiringen ophørte, før temperaturerne blev ændret, konkluderede vi, at manglen på yderligere spiring før vinteren ikke var drevet af temperatur (A3). I modsætning hertil syntes spiring af meget af A2-gruppen kun at blive standset, når temperaturerne blev reduceret til 10/5 C. C., Klynge B2 indeholdt yderligere fire arter, der udviste betydelig spiring tidligt i eksperimentet og fortsatte med at opnå en relativt lille andel af deres samlede spiring, efter at temperaturerne blev returneret til over 5.C (figur 3B).
sammenlignet med den relativt mindre variation blandt arter inden for klynger A1 og B1, viser de forskudte klynger også meget større differentiering blandt arter. De enkelte kurver for alle arter er vist i Supplerende Tal 1i–vii. For mere specifikke detaljer om spiring strategi, se Appendiks B i Supplerende Materiale.,
Økologiske Korrelater til Spiring Strategi
Af alle de økologiske korrelerer, at vi overvejet, kun dem, der er direkte forbundet med frø træk var signifikant korreleret med spireevne strategi, uanset om vi vurderet sammenhængen på de to store klynger (udsat vs. umiddelbar spiring), eller på niveau tre eller fire klynger, eller om vi tegnede sig for fylogenetiske struktur eller ej (Tabel 2)., Arter med udskudt spiring havde tungere frø og var mere tilbøjelige til at være endospermiske sammenlignet med arter, der spirede straks (figur 4). Arter med forskudt spiring var mellemliggende i frømasse og endospermi. Imidlertid var ikke alle frøegenskaber korreleret med spiringsstrategi: der var ingen sammenhæng mellem lyskrav til frøspiring og strategi. I modsætning til vores forventninger var arter med højere specifikt bladområde (potentielt tegn på højere vækstrater) ikke mere tilbøjelige til at udvise øjeblikkelig spiring., Ligeledes var arter med højere gennemsnitlig højde eller mindre elevationsområder ikke mere tilbøjelige til at udvise udskudt spiring.

Tabel 2. Forekomst af fylogenetiske struktur i, og korrelater af, spiring strategi.
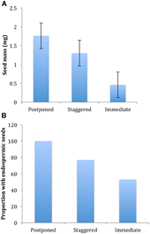
Figur 4., Forskelle i (A) gennemsnitlig frømasse og (B) andel af arter med endospermiske frø blandt arter med udskudt (klynge A1), forskudt (kombination af klynge A2, A3 og B2) og umiddelbar spiring (klynge B1) strategier. Forskelle mellem strategier er signifikante ved P 0.0 0,05, se tabel 2.
Diskussion
Spiring i alpine levesteder, der traditionelt anses for at være miljømæssigt kontrolleret, med vinter, sne isolerende frø mod potentielle spiring stikord og rendering vækstdvale unødvendige (Billings og Mooney, 1968)., Vores resultater understøtter dog nyere bevis for, at dvalemekanismer er almindelige blandt alpine arter både i Australien (Hoyle et al., 2013; Sommerville et al., 2013) og andre steder i verden (Cavieres og Arroyo, 2000; Schutz, 2002; Shimono og Kudo, 2005; Mondoni et al., 2009; Sch .ienbacher et al., 2011)., Vækstdvale ville synes at spille en væsentlig rolle i at kontrollere in situ timing af spiring af mange Australske alpine arter, der handler om at forsinke, udskyde eller nedsætte den hastighed, hvormed spiring og dermed potentielt overdragelse af risiko-forcernes regenerering strategier i en barsk og variabel miljø. Måske mest slående i vores spiring resultater var andelen af arter, for hvilke spiring strategi varieret inden for en frø samling for en given art, der tyder på, at både sovende og ikke-sovende egenskaber blev udstillet i samme frø samling af disse arter., Især blev spiringsstrategier generelt stærkt bevaret inden for familier og også korreleret med elementer af frøanatomi: masse og endospermi. Spiringsstrategier var imidlertid uafhængige af andre økologisk betydningsfulde funktionelle træk ved den modne plante.
Variation i Spiringsstrategi
frø af næsten halvdelen af de undersøgte arter syntes ikke at spire, før de blev udsat for en kølig, våd periode (konstant 5.C), hvilket antydede, at kold stratificering lindrede en fysiologisk sovemekanisme., Udsættelse af spiring til det følgende forår kan gøre det muligt for frøplanter at undgå at etablere sig over eller før den hårde vinter, samtidig med at den korte kommende vækstsæson optimeres. I modsætning til den opportunistiske spiring udstillet af de arter, der spirede straks kan give en selektiv fordel, når risikoen for vinteren sætteplante dødeligheden er lav, ved at sætte planter til at blomstre tidligere, det følgende forår, eller i en større størrelse (Donohue, 2002).,
Til dato har der været lidt offentliggjorte dokumentation af intra-specifikke variation i vækstdvale mekanismer i alpine frø samlinger, som ville kunne forklare den forskudte spiring strategier, der er observeret her. Spiring forskudt over tid kan også forklares ved forskellige niveauer af frømodning blandt individer i befolkningen på indsamlingstidspunktet og/eller kunne afspejle en dimorf strategi inden for en enkelt plante forbundet med position på planten eller tidspunktet for udvikling.,
evolutionær bet-hedging fremkaldes ofte for at forklare den diversificerede strategi i frøspiringskarakteristika (Simons, 2011), som det demonstreres af samlingerne med forskudt spiring. Hvis en diversificeret strategi imidlertid resulterer i både øget gennemsnitlig kondition og en reduktion i variansen (f.eks. ved at reducere konkurrencen mellem søskende), udgør den ikke væddemålssikring. Ligeledes, hvis den tilsyneladende diversificering afspejler forskelle mellem, ikke inden for enkeltpersoner, så er det ikke bet-hedging (Starrfelt and Kokko, 2012)., Undersøgelser alpine sætteplante spredning mønstre og tolerance over for frost og sne smelter, parret med undersøgelser af determinanter for spiring strategi variation inden for arter, der kan hjælpe med at forklare spiring phenology af de arter, der udviser en forskudt strategi, og kan oplyse, om de er tilbøjelige til at repræsentere bet-hedging strategier.,
økologiske korrelater af spiring strategi
vores undersøgelse af forholdet mellem spiring strategi og andre økologisk vigtige træk viste, at spiring træk var korreleret med andre frø træk, men ikke med vegetative træk. Sommerville et al. (2013) fandt, at Australske alpine urter med ikke-endospermiske frø var mere tilbøjelige til at være ikke-sovende ved spredning. Vores arbejde understøtter dette fund og indikerer yderligere, at mindre frø er mere tilbøjelige til at være blandt de umiddelbare spirere. Selvom Sommerville et al., (2013) stillede ikke hvorfor endospermi kan være forbundet med dvaletid, vi foreslår, at placering af reserver i cotyledoner kan forbedre den tidlige vækstrate og etablering I umiddelbare spirende arter. Vores resultater indikerede ikke nogen sammenhæng mellem lysbehov og dvaletilstand, selvom lyskravene varierer mellem arter. Der er en række andre frøegenskaber, der buffer udryddelsesrisiko i variable miljøer, for eksempel frøspredningsmekanismer og frø levetid i jordfrøbanken (se Tielborger et al., 2012 for en gennemgang)., På dette stadium kan vi ikke sige, om disse træk ville vise korrelationer med spiringsstrategi eller ej.
Vi spurgte endvidere, om spiringsstrategi ville være korreleret med økologisk vigtige ikke-reproduktive træk, såsom specifikt bladområde eller plantehøjde (1998estoby, 1998; .right et al., 2004). I særdeleshed vi postulerede, at den umiddelbare spirende arter kan vise værdierne af vejledende hurtigere vækstrater (ved hjælp af SLA som en proxy), som kan være nødvendige for etablering i den korte periode mellem spredning og starten af vinteren., Vi fandt dog ingen sådanne korrelationer. Hvorvidt fysiologiske og vækstegenskaber hos frøplanter i tidlige etableringsstadier kan vise forskellige mønstre end voksne vegetative træk er et spørgsmål, der er værd at overveje, men uden for rammerne af det nuværende arbejde.
Endelig, vi har undersøgt, om arter, der fra højere niveauer, hvor vækstsæsoner er generelt kortere kan være mere tilbøjelige til at udstille vækstdvale, men der var ingen association mellem enten højde interval, indsamling af stigninger, eller den gennemsnitlige højde af indsamling for arten som helhed og spiring strategi., Sammen viser disse resultater, at mens udviklingen af spiringsstrategi er tæt knyttet til økologisk vigtige frøegenskaber, omfatter den en stort set uafhængig akse til vegetative eller fordelende egenskaber.
Konsekvenser for Alpine Anlæg Fællesskaber i et Skiftende Klima
den Australske alpine økosystemer, som alpine områder rundt omkring i verden, er truet af klimaforandringer kombineret med ændringer i brand frekvens, arealanvendelse mønstre, tilstrømning af invasive arter, og konsekvenserne af øget menneskelig visitation., Opvarmning i forbindelse med klimaændringer forekommer hurtigere over trælinjen end i lavere højder, og alpine områder forventes fortsat at opleve opvarmning over gennemsnittet i fremtiden (Kullman, 2004). Kapaciteten til fortsat regenerering via frø under nye forhold vil sandsynligvis spille en væsentlig rolle i Alpine planters reaktion på klimaet og dermed forbundne ændringer. Desuden vil ændrede forstyrrelsesordninger sandsynligvis kræve en øget rolle for restaurering og rehabilitering i regionen., Den indsigt, der er fastsat her, er derfor af afgørende betydning for bevarelsen og forvaltningen alpine systemer under forandring, og de kan hjælpe med at informere vores forudsigelser af, hvordan undersøgelsen arter kan blive påvirket, danne grundlag for frøformering planer for disse arter, og bruges til at guide den fremtidige undersøgelse af alpine frøspiring strategier med identificerbare bevarelse og forvaltning resultater.,
temperaturændringer vil sandsynligvis påvirke frøenes sovestatus både før og efter spredning, hvilket følgelig ændrer og/eller forstyrrer spiringsstrategier og resulterer i skift i den nuværende spirefænologi (Hoyle et al., 2008b). For alpine planter, klima opvarmning kan føre til et skift fra forår til efterår opstår, primært drevet af ændringer i frø vækstdvale status og resulterer i betydelige konsekvenser for de arter, der i øjeblikket er tilpasset til fremkomsten i foråret (Mondoni et al., 2012)., Yderligere forskning i, hvordan ændringer i temperaturer, der frø erfaring post-spredning, reduceret varighed af sne, og øget frekvens af ekstreme temperatur begivenheder vil påvirke spiringen er nødvendige for at forudsige, hvordan arter med vækstdvale vil reagere på klimaændringer. For de alpine arter, der spirer i efteråret, før frost og sne, vores resultater rejser spørgsmålet om, hvordan frøplanter af disse arter vil kunne klare den forventede reduktion i nedbør og øget frekvens af a-sæsonåben frost begivenheder.,
endelig rejser vores resultater vigtige spørgsmål vedrørende variation inden for arter, især i forbindelse med at forudsige virkningen af klimaændringer eller vælge hvilke frø der skal bruges til restaurering. Spiring strategier kan variere inden for de enkelte arter, afhængigt af befolkningen, blomstringstid, frø masse, forældrenes investeringer, klima, højde og/eller moderens miljø og denne variation kan skyldes genetiske, epigenetiske, og/eller miljømæssige faktorer (Cochrane et al., 2015)., Stigende højde tegner sig for variation i germinability og/eller vækstdvale status af flere alpine arter, (Billings og Mooney, 1968; Dorne, 1981; Cavieres og Arroyo, 2000), men se (Hoyle et al., 2014). Reduceret dvalestatus kan skyldes tidlig afslutning af frøudvikling som følge af miljøbelastninger (Steadman et al., 2006; Hoyle et al., 2008b)., Undersøgelse af variation i spiringsstrategier inden for arter kan afsløre potentialet for arter til at klare og tilpasse sig ændringer i klimaet, dermed, spiller en vigtig rolle i bestemmelsen af fremtidig overlevelse og artsfordelinger og giver vigtig indsigt til forvaltning og bevaring.
konklusioner
vores tilgang har afsløret udbredt forekomst af dvaletilstand i den australske alpine flora og demonstrerer, at spiringsstrategier varierer inden for en frøopsamling såvel som mellem samlinger., Yderligere, vi har vist, at mens dvalestrategi er fylogenetisk konserveret og korreleret med frømasse og endospermi, det er stort set uafhængigt af vegetative træk og rækkevidde. Mens man ikke kan ekstrapolere direkte fra vores resultater, hvordan klima og tilknyttede ændringer vil påvirke alpine flora, kan disse resultater danne grundlag for udformning af formeringsplaner og yderligere eksperimentering af virkningen af ændrede klimatiske regimer., Vi går især ind for yderligere forskning i forståelse af årsagerne til variation i spiringsstrategien inden for arter og den rolle, dette kan spille i arternes evne til at modstå forudsagte klimaændringer.
data tilgængelighed
fuld spiring data og træk database vil blive indgivet på DRYAD og med Atlas of Living Australia.
Erklæring om interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.,
anerkendelser
Vi anerkender finansiering fra Australian Research Council (LP0991593) og Australian National Botanic Garden og vennerne deraf. Vi takker J. McAuliffe, S. Fethers og S. La .atsch fra ANBG. Vi takker også M. Hollo .ay-Phillips, M. Cardillo og S. Cunningham for input til tidligere versioner af manuskriptet og for analytisk rådgivning og S. Venn for adgang til trækdata.
supplerende materiale
Foley, M. E. (1992). Virkningen af opløselige sukkerarter og gibberellic-syre ved brud på dvale af udskårne vilde havre (Avena fatua) embryoner., Ukrudtsscanning. 40, 208–214.
Google Scholar
Harden, G. J. (1990). Flora of New South Wales. Sydney, NS.: Royal Botanic Gardens.
Google Scholar
International Seed Testing Association. (2003). Arbejder ark på Tetra .olium test. Bassersdorf: ISTA.
Kullman, L. (2004). Den skiftende ansigt af den alpine verden. Glob. Skift Ne .sl. 57, 12–14.
Google Scholar
Skriv et svar