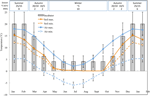
Figura 1. Datos mensuales de temperatura máxima y mínima del aire y del suelo en los Alpes Australianos (media ± STDEV). Las temperaturas del aire son promedios de la base de datos BOM, las temperaturas del suelo se midieron con iButtons (ver métodos). Las barras indican las temperaturas de la incubadora experimental (media ± STDEV)., La época equivalente del año (estación), el régimen de temperatura de la incubadora (°C, 12/12 h, claro/oscuro) y la duración del tratamiento (semanas) según se indica en el panel superior.
con el fin de descubrir condiciones de temperatura cercanas a las que las semillas experimentan después de la dispersión, se recolectaron datos diarios de temperatura máxima y mínima del suelo dentro del sitio de estudio (Seaman’s Hut; 36.27°S y 148.17°E, 2030 m s. n.m.) utilizando registradores de datos ibutton (n = 10, Embedded Data Systems, EE. de vegetación del 17 de enero al 17 de diciembre de 2012., Colocamos taladores a 4 cm para evitar perturbaciones superficiales pero aún así ser representativos de las condiciones a las que las semillas están expuestas en el suelo; esta profundidad es intermedia a lo que se ha utilizado en ejercicios previos de monitoreo de la temperatura del suelo alpino (Scherrer y Körner, 2011; Pauli et al., 2015). Como se sospecha, la temperatura media del suelo no cayó por debajo del punto de congelación durante el invierno (Figura 1). En verano, se sabe que la temperatura del suelo bajo la vegetación rastrea la temperatura ambiente, mientras que las temperaturas del suelo desnudo excederán el ambiente en hasta 15°C (registros inéditos del servicio de conservación del suelo, 1960s–1970s).,
recolección y germinación de semillas
se recolectaron semillas maduras de 54 especies de 16 familias y 37 géneros entre enero y abril de 2009, 2010 y 2011 (Ver Tabla 1 para nombres completos y autoridades. Vouchers were lodged at the Australian National Herbarium, Canberra). En total, la especie representaba más de una cuarta parte de la flora angiosperma Australiana encontrada en regiones alpinas (Costin et al., 2000, Ver Tabla 1), aunque muchas de estas especies también se extienden por debajo del límite arbóreo arbóreo., La viabilidad de todas las colecciones se estimó antes de la siembra en condiciones experimentales de germinación utilizando la técnica de tinción de cloruro de tetrazolio (TZ) (Asociación Internacional de pruebas de semillas, 2003). Para más detalles sobre la recopilación y el procesamiento, véase el Apéndice A en material suplementario.
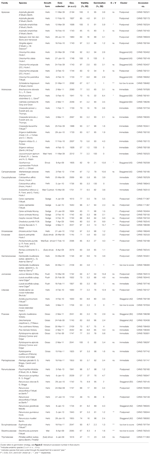
la Tabla 1. Estudio de la especie, familia y autoridad, forma de crecimiento y detalles de las colecciones, porcentaje de viabilidad de la semilla (TZ-estimado, ± s.e.), porcentaje final de germinación alcanzada (±s.e.,), diferencia estadística entre viabilidad y germinación (G < V, probado solo en casos donde la germinación fue menor que la viabilidad).
la fenología de la germinación de semillas se investigó imitando los regímenes de temperatura que las semillas alpinas experimentan in situ, después de la dispersión, en una progresión artificialmente acortada de las estaciones. Las semillas fueron absorbidas durante todo el experimento, por lo que los resultados indican un potencial de germinación cuando el agua no es limitante., Los datos de temperatura del aire y del suelo (Figura 1) se utilizaron para guiar y validar los regímenes de temperatura de las incubadoras, aunque las contraindicaciones logísticas del trabajo con incubadoras impidieron la incorporación de las fluctuaciones inherentes a las condiciones naturales. Las pruebas de germinación utilizaron ocho réplicas de 25 semillas por colección, sembradas en placas de Petri de plástico de 9 cm de diámetro que contenían 1% de agar de agua simple. Las placas de Petri fueron selladas con Parafilm para evitar la desecación del agar, antes de ser colocadas en incubadoras de germinación (Thermoline Scientific, Melbourne, NSW, Australia)., La mitad de los platos se envolvieron en papel de aluminio para excluir la luz y proporcionar una indicación de los requisitos de luz para la germinación. Cada una de las cuatro réplicas por tratamiento de luz por colección se colocó en un estante diferente dentro de la misma incubadora y la ubicación dentro de ese estante se reorganizó semanalmente. Todas las réplicas de cada colección se movieron a través de los siguientes termoperiodos simulados: 8 semanas a 25/15°C (Temperatura día/noche) → 2 semanas a 20/10°c → 2 semanas a 10/5°c → 10 semanas a constante 4-5°C → 2 semanas a 10/5°c → 2 semanas a 20/10°c → 6 semanas a 25/15°C., Por lo tanto, un ciclo de todo el experimento imitó temperaturas que recuerdan al verano, principios de otoño, finales de otoño, invierno, principios de primavera, finales de primavera y verano, y duró 32 semanas en total. El año abreviado se consideró razonable porque las accesiones generalmente habían dejado de germinar en un tratamiento dado antes de ser cambiadas a la siguiente «estación»., Un fotoperiodo claro/oscuro de 12/12 h fue proporcionado por tubos fluorescentes (> 90 µmol m-2 s-1 en el Centro de cada estante), y en los regímenes de temperatura de primavera, verano y otoño la luz coincidió con el período de temperatura cálida del día. La germinación, definida como la aparición de radículas por más de 1 mm, se anotó cada 7 días, y las semillas germinadas y las plántulas se retiraron posteriormente de los platos. La germinación de las placas envueltas en papel de aluminio se evaluó como se indicó anteriormente, pero con muy poca luz verde.,
Después de un ciclo experimental (32 semanas) la mayoría de las colecciones fueron terminadas (ver información suplementaria para excepciones), habiendo exhibido cero germinación durante al menos 2 semanas en este momento. Después de la terminación, todas las semillas restantes intactas fueron diseccionadas con un bisturí bajo un microscopio. Las semillas con un endospermo firme y fresco y el embrión se consideraron viables y las semillas vacías de un embrión se dedujeron del total al calcular el porcentaje de germinación para cada colección., Se calculó un índice de los requerimientos de luz para germinación para cada especie como una fracción de germinación en el tratamiento oscuro en el momento en que se alcanzaron percentiles de germinación acumulativos de 50 y 95% en el tratamiento alterno claro/oscuro (Ver más adelante).
correlatos ecológicos de la estrategia de germinación
se compiló una base de datos de rasgos funcionales y medidas geográficas de especies a partir de datos publicados para evaluar correlaciones con la estrategia de germinación., La base de datos incluyó los siguientes rasgos continuos: masa media de la semilla, requerimiento de luz para la germinación (ver arriba), área foliar específica (área fresca/masa seca), área foliar individual, peso seco individual de la hoja y altura en la madurez. Las elevaciones mínimas, medias y máximas de recolección para la especie se determinaron a partir de todos los registros de herbario en el Herbario Virtual australiano, y la elevación a la que se realizaron las colecciones de semillas específicas utilizadas en el estudio también se incluyó como rasgos continuos. La forma de crecimiento se clasificó como una de tres categorías: graminoide, hierba o arbusto., También incluimos un rasgo binario: si las semillas eran endospermicas o no.
los datos del rasgo fueron recolectados de una variedad de fuentes incluyendo el Herbario Virtual Australiano (http://avh.chah.org.au/), el registro de información de especímenes del Herbario Nacional Australiano (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Alpine Flora (Costin et al., 2000), The New South Wales Flora (Harden, 1990), and the comparative internal morphology of seeds (Martin, 1946). La masa de semilla y el área foliar específica se obtuvieron de colecciones de campo individuales utilizadas en los ensayos de germinación., La masa de semillas se obtuvo pesando cuatro lotes de 10 semillas. El SLA se determinó en base a cinco hojas completamente expandidas de individuos separados por especie. Estos fueron recolectados en el campo, escaneados en un escáner de cama plana, secados a 60 ° C a Peso constante y ponderados a 0.000 g más cercanos. cuando no se disponía de semilla suficiente o no se disponía de área foliar específica, se obtuvieron datos de otras colecciones de ANBG de la misma especie o de otras fuentes de datos (S. Venn, datos no publicados)., Se determinó si una semilla era endospermica o no a partir de dibujos de la semilla en los registros del Banco de semillas de ANGB en consulta con el personal del Banco de semillas de ANBG. La base de datos completa y las fuentes están disponibles a petición de los autores.
análisis de datos
Los patrones de germinación fueron evaluados visualmente y utilizando un análisis de cluster en PC-ORD Para Windows, versión 6 (MJM Software Design, Gleneden Beach, OR, USA). El análisis utilizó una matriz de distancia euclidiana y un algoritmo de unión del vecino más cercano., Los puntos de datos para el análisis de conglomerados fueron el tiempo (semanas) hasta los percentiles de germinación acumulativos específicos (t20, t40, t50, t60, t80 y t95%) para cada colección (promediados en los 4 platos). Por lo tanto, una colección que germinó temprano en el experimento tendría tiempos cortos a percentiles hasta t80%, por ejemplo, mientras que una colección que pospuso la germinación hasta que las temperaturas aumentaran después del invierno simulado tendría tiempos largos a estos percentiles., También se podrían detectar patrones de germinación más complejos, como germinación lenta y constante o ráfagas de germinación, y se indicarían por tiempos espaciados uniformemente o tiempos agrupados al comienzo y al final del experimento, respectivamente. Este método fue utilizado como una forma de imponer una evaluación objetiva del patrón de germinación, independiente de la temperatura per se.,
se realizó un análisis de varianza unidireccional (ANOVA) para las placas replicadas de cada colección para evaluar si había una diferencia significativa entre el porcentaje de viabilidad (determinado por la prueba TZ) y el porcentaje final de germinación. Se utilizaron otros modelos lineales generales para evaluar si los racimos resultantes del análisis de racimos diferían en viabilidad o porcentaje de germinación final.
Las asociaciones entre los rasgos ecológicos y los clusters de la estrategia de germinación se probaron de dos maneras., En primer lugar, probamos las diferencias medias entre los clústeres utilizando ANOVA con clústeres como efecto fijo y correlatos ecológicos como variables de respuesta, o utilizando pruebas g replicadas para la endosperma y la forma de crecimiento. En segundo lugar, debido a que es muy probable que la germinación y otros rasgos funcionales estén influenciados por la historia evolutiva, evaluamos estas correlaciones utilizando regresiones filogenéticas. Se construyó una filogenia para las especies de estudio utilizando Phylocom Phylomatic y el bladj packing en R (Webb et al., 2008; R Core Team, 2013)., Se evaluó la presencia de estructura filogenética en variables utilizando el phylosig o phylo.D (discrete or binary variables) función del paquete Phytools en R (Revell, 2012). Las regresiones filogenéticas se realizaron utilizando la función pgls en Caper (Cape et al., 2013). Los análisis se llevaron a cabo de una manera estructurada que refleja la agrupación revelada en PC-Ord (véase más adelante)., Inicialmente incluimos cuatro estrategias de germinación (pospuesta, inmediata y escalonada dentro de cada una de las anteriores), luego tres (agrupando todas las especies escalonadas y comparándolas con pospuesta e intermedia) y finalmente evaluamos las diferencias entre las dos estrategias más amplias, pospuesta e intermedia.
resultados
viabilidad de la semilla en comparación con la germinación final
En general, la viabilidad de la semilla y el porcentaje de germinación final fueron altos. La viabilidad media estimada por TZ en todas las colecciones fue de 80 ± 1.,7%, con más de dos tercios de las colecciones que exhiben más del 75% de viabilidad (Tabla 1). No hubo diferencia en la calidad de las colecciones a lo largo de los 3 años (la viabilidad media en 2009, 2010 y 2011 fue de 79 ± 2.8, 80 ± 2.7 y 81 ± 3.7%, respectivamente), lo que indica que el banco de semillas recolectadas en 2009 no disminuyó su viabilidad.
Se logró germinación que excedió o no fue significativamente menor a la viabilidad estimada por TZ para 45 de las 54 especies (ANOVA unidireccional: P>0.05, Tabla 1)., De las nueve especies restantes, la germinación final de cuatro especies fue muy baja (< 5%), a pesar de la buena viabilidad (ca. 55-94%), por lo que no pudimos sacar ninguna conclusión sobre las estrategias de germinación de estas especies. La falta de germinación podría reflejar que nuestros tratamientos no aliviaron la latencia o que las semillas no estaban completamente maduras en la recolección. Las pruebas de corte al final del experimento indicaron que, en todos los casos, la mayoría de las semillas no germinadas contenían un embrión sano y embebido., Curiosamente, 34 especies lograron una germinación mayor que la viabilidad estimada por TZ (Tabla 1), lo que sugiere que la prueba de TZ comúnmente subestimó la viabilidad de la semilla.
Clusters de germinación
el análisis de clusters reveló dos clusters principales (A y B), con subestructura en cada uno (Figura 2). Para la mayoría de las especies (n = 21 especies de 10 familias), la germinación se pospuso hasta durante un período de temperaturas invernales (5°C día/noche) o hasta que las temperaturas se elevaron a 10/5°C después del invierno (A1 y A1:invierno, Figuras 2, 3a)., Alternativamente, la germinación comenzó inmediatamente a 25/15 ° C (n = 17 especies, seis familias, vistas en el cluster B1, Figuras 2, 3b). Para la mayoría de estas especies, la tasa de germinación fue rápida; el 50% de la germinación total (t50) ocurrió en menos de 3 semanas (figura suplementaria 1).
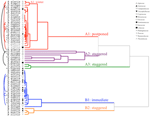
la Figura 2. Dendrograma de análisis de conglomerados basado en el tiempo hasta los percentiles de germinación acumulativos (20, 40, 50, 60, 80, y 95%). Especie indicada por la inicial del género y la primera parte del nombre de la especie. Los asteriscos marcan las colecciones que se almacenaron antes de las pruebas., Los arcos finos conectan colecciones duplicadas para comparar el efecto de la banca en la germinación. Los símbolos denotan diferentes familias (ver leyenda). Las colecciones mostradas en texto gris son aquellas que fueron expuestas a dos ciclos del experimento.
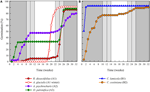
la Figura 3. Porcentaje acumulado de germinación de una especie representativa de cada racimo de germinación. (A) grupo a, especies que pospusieron la germinación, (B) Grupo B, especies que germinaron inmediatamente., Tenga en cuenta que los patrones de germinación escalonados están anidados dentro de los grupos anteriores. Los regímenes de temperatura de la incubadora están representados por sombreado (ver Figura 1).
dentro de cada una de estas clases amplias, sin embargo, había subgrupos claros que reflejaban una germinación escalonada a lo largo del tiempo, es decir, una proporción del lote de semillas germinó antes de la exposición a las temperaturas invernales, mientras que el resto germinó después del invierno. La germinación escalonada ocurrió en 13 especies de seis familias Y ocurrió principalmente cuando las temperaturas diurnas excedieron los 10°C (figuras 3A,B)., Estos germinadores escalonados se dividieron en tres grupos en el análisis de conglomerados. El racimo A2 con siete colecciones mostró una germinación sustancial tanto antes como después del período de temperatura fría. Donde la germinación cesó antes de que se cambiaran las temperaturas, concluimos que la falta de germinación antes del invierno no fue impulsada por la temperatura (A3). En contraste, la germinación de gran parte del grupo A2 pareció detenerse solo cuando las temperaturas se redujeron a 10/5°C., El grupo B2 contenía otras cuatro especies que exhibieron una germinación sustancial al principio del experimento y lograron una proporción relativamente pequeña de su germinación total después de que las temperaturas volvieran a estar por encima de los 5°C (figura 3b).
en comparación con la variación relativamente menor entre las especies dentro de los grupos A1 y B1, los grupos escalonados también muestran una diferenciación mucho mayor entre las especies. Las curvas individuales para todas las especies se muestran en las figuras suplementarias 1i-vii. para detalles más específicos sobre la estrategia de germinación, véase el Apéndice B en material suplementario.,
correlatos ecológicos de la estrategia de germinación
de todos los correlatos ecológicos que consideramos, solo aquellos directamente asociados con los rasgos de la semilla se correlacionaron significativamente con la estrategia de germinación, independientemente de si evaluamos la correlación a nivel de dos clústeres principales (germinación pospuesta vs.inmediata), o a nivel de tres o cuatro clústeres, o si explicamos la estructura filogenética o no (Tabla 2)., Las especies con germinación pospuesta tenían semillas más pesadas y tenían más probabilidades de ser endospermicas en comparación con las de las especies que germinaron inmediatamente (Figura 4). Las especies con germinación escalonada fueron intermedias en masa de semillas y endospermas. Sin embargo, no todas las características de la semilla se correlacionaron con la estrategia de germinación: no hubo correlación entre los requisitos de luz para la germinación de la semilla y la estrategia. Contrariamente a nuestras expectativas, las especies con mayor área foliar específica (potencialmente indicativa de tasas de crecimiento más altas) no tenían más probabilidades de exhibir germinación inmediata., Del mismo modo, las especies con elevación promedio más alta o rangos de elevación más pequeños no tuvieron más probabilidades de presentar una germinación pospuesta.

la Tabla 2. Incidence of phylogenetic structure in, and correlates of, germination strategy.
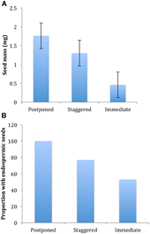
la Figura 4., Diferencias en (A) masa media de semillas y (B) proporción de especies con semillas endospermicas entre las especies con estrategias de germinación pospuesta (cluster A1), escalonada (combinación de cluster A2, A3 y B2) e inmediata (cluster B1). Las diferencias entre las estrategias son significativas en p ≤ 0,05, Ver Tabla 2.
discusión
La germinación en hábitats alpinos se consideró históricamente controlada ambientalmente, con la nieve de invierno aislando las semillas contra posibles señales de germinación y haciendo innecesaria la latencia (Billings y Mooney, 1968)., Sin embargo, nuestros resultados apoyan la evidencia más reciente de que los mecanismos de latencia son comunes entre las especies alpinas tanto en Australia (Hoyle et al., 2013; Sommerville et al., 2013) y en otras partes del mundo (Cavieres y Arroyo, 2000; Schutz, 2002; Shimono y Kudo, 2005; Mondoni et al., 2009; Schwienbacher et al., 2011)., La latencia parece desempeñar un papel importante en el control del Tiempo in situ de germinación de muchas especies alpinas australianas, actuando para retrasar, posponer o ralentizar la tasa de germinación y, por lo tanto, potencialmente confiriendo estrategias de regeneración reacias al riesgo en un entorno duro y variable. Quizás lo más sorprendente de nuestros resultados de germinación fue la proporción de especies para las que la estrategia de germinación varió dentro de una colección de semillas para una especie dada, lo que sugiere que tanto las características latentes como las no latentes se exhibieron dentro de la misma colección de semillas de estas especies., Notablemente, las estrategias de germinación fueron generalmente altamente conservadas dentro de las familias y también se correlacionaron con elementos de la anatomía de la semilla: masa y endosperma. Las estrategias de germinación fueron, sin embargo, independientes de otros rasgos funcionales ecológicamente significativos de la planta madura.
variación en la estrategia de germinación
Las semillas de casi la mitad de las especies estudiadas no pudieron germinar hasta que fueron expuestas a un período fresco y húmedo (constante de 5°C), lo que sugiere que la estratificación en frío alivió un mecanismo fisiológico de latencia., Posponer la germinación hasta la primavera siguiente puede permitir que las plántulas eviten establecerse durante o antes del duro invierno, al tiempo que optimiza la corta temporada de crecimiento. En contraste, la germinación oportunista exhibida por las especies que germinaron inmediatamente puede proporcionar una ventaja selectiva cuando el riesgo de mortalidad de plántulas en invierno es bajo, al permitir que las plantas florezcan más temprano en la primavera siguiente o en un tamaño mayor (Donohue, 2002).,
hasta la fecha, ha habido poca evidencia publicada de variabilidad intraespecífica en los mecanismos de latencia dentro de las colecciones de semillas alpinas, como explicaría las estrategias de germinación escalonadas observadas aquí. La germinación escalonada a lo largo del tiempo también puede explicarse por los diferentes niveles de madurez de la semilla entre los individuos de la población en el momento de la recolección, y/o podría reflejar una estrategia dimórfica dentro de una sola planta asociada con la posición en la planta o el momento del desarrollo.,
La cobertura de apuesta evolutiva se evoca a menudo para explicar la estrategia diversificada en las características de germinación de semillas (Simons, 2011), como la demostrada por las colecciones con germinación escalonada. Sin embargo, si una estrategia diversificada resulta tanto en un aumento de la aptitud promedio como en una reducción de la varianza (por ejemplo, al reducir la competencia entre hermanos), entonces no constituye cobertura de apuestas. Del mismo modo, si la diversificación aparente refleja diferencias entre individuos, no dentro de ellos, entonces no es una cobertura de apuesta (Starrfelt y Kokko, 2012)., Las investigaciones sobre los patrones de dispersión de plántulas alpinas y la tolerancia a las heladas y el derretimiento de la nieve, junto con estudios de los determinantes de la variación de la estrategia de germinación dentro de las especies, pueden ayudar a explicar la fenología de germinación de las especies que exhiben una estrategia escalonada y pueden indicar si es probable que representen estrategias de cobertura bet.,
correlatos ecológicos de la estrategia de germinación
nuestro examen de la relación entre la estrategia de germinación y otros rasgos ecológicamente importantes demostró que los rasgos de germinación estaban correlacionados con otros rasgos de semilla pero no con rasgos vegetativos. Sommerville et al. (2013) encontraron que las hierbas alpinas australianas con semillas no endospermicas tenían más probabilidades de no estar latentes en la dispersión. Nuestro trabajo apoya ese hallazgo e indica además que las semillas más pequeñas tienen más probabilidades de estar entre los germinadores inmediatos. Aunque Sommerville et al., (2013) no postularon por qué la endosperma puede estar asociada con la latencia, sugerimos que la colocación de reservas en cotiledones puede mejorar la tasa de crecimiento temprano y el establecimiento En especies de germinación inmediata. Nuestros resultados no indicaron ninguna correlación entre el requerimiento de luz y la latencia, aunque los requerimientos de luz variaron entre las especies. Hay una variedad de otros rasgos de semillas que amortiguan el riesgo de extinción en ambientes variables, por ejemplo los mecanismos de dispersión de semillas y la longevidad de las semillas en el banco de semillas del suelo (ver Tielborger et al., 2012 for a review)., En esta etapa no podemos decir si esos rasgos mostrarían correlaciones con la estrategia de germinación o no.
Además, preguntamos si la estrategia de germinación estaría correlacionada con rasgos no reproductivos ecológicamente importantes, como el área foliar específica o la altura de la planta (Westoby, 1998; Wright et al., 2004). En particular, planteamos que las especies de germinación inmediata podrían mostrar valores indicativos de tasas de crecimiento más rápidas (utilizando SLA como un proxy) como podría ser necesario para el establecimiento En el corto período entre la dispersión y el inicio del invierno., Sin embargo, no encontramos tales correlaciones. Si los rasgos fisiológicos y de crecimiento de las plántulas en las primeras etapas de establecimiento podrían mostrar patrones diferentes que los rasgos vegetativos adultos es una cuestión digna de consideración, pero más allá del alcance del presente trabajo.
finalmente, se examinó si las especies de elevaciones más altas donde las estaciones de crecimiento son generalmente más cortas podrían tener más probabilidades de exhibir latencia, pero no hubo asociación entre el rango de elevación, las elevaciones de la colección o la elevación promedio de la colección para la especie en su conjunto y la estrategia de germinación., En conjunto, estos resultados indican que, si bien la evolución de la estrategia de germinación está estrechamente vinculada a rasgos de semillas ecológicamente importantes, comprende un eje en gran medida independiente de las características vegetativas o distributivas.
Implications for Alpine Plant Communities in a Changing Climate
Australian alpine ecosystems, like alpine areas around the world, are under threat from climate change combined with changes in fire frequency, land use patterns, influx of invasive species, and the impacts of increased human visitation., El calentamiento asociado con el cambio climático está ocurriendo más rápidamente por encima del límite arbóreo superior que en elevaciones más bajas, y se prevé que las áreas alpinas continúen experimentando un calentamiento por encima del promedio en el futuro (Kullman, 2004). Es probable que la capacidad de regeneración continua a través de semillas en condiciones nuevas desempeñe un papel importante en la respuesta de las plantas alpinas al clima y los cambios asociados. Además, es probable que los regímenes de perturbación alterados requieran un mayor papel de restauración y Rehabilitación en la región., Por lo tanto, los conocimientos proporcionados aquí son cruciales para la conservación y el manejo de los sistemas alpinos en cambio; pueden ayudar a informar nuestras predicciones de cómo las especies de estudio pueden verse afectadas, formar la base de los planes de propagación de semillas para estas especies y ser utilizados para guiar la investigación futura sobre las estrategias de germinación de semillas alpinas con resultados identificables de conservación y manejo.,
Es probable que los cambios de temperatura afecten el estado de latencia de las semillas tanto antes como después de la dispersión, alterando y/o interrumpiendo las estrategias de germinación y dando como resultado cambios en la fenología de germinación actual (Hoyle et al., 2008b). Para las plantas alpinas, el calentamiento del clima puede conducir a un cambio de la emergencia de primavera a otoño, impulsado principalmente por cambios en el estado de latencia de las semillas y dando lugar a importantes implicaciones para las especies actualmente adaptadas a la emergencia en primavera (Mondoni et al., 2012)., Se requieren más investigaciones sobre cómo los cambios en las temperaturas que experimentan las semillas después de la dispersión, la reducción de la duración de la cubierta de nieve y el aumento de la frecuencia de eventos de temperatura extrema afectarán la germinación para predecir cómo las especies con latencia responderán al cambio climático. Para aquellas especies alpinas que germinan en otoño, antes de las heladas y las nevadas, nuestros resultados plantean la pregunta de cómo las plántulas de estas especies harán frente a la reducción prevista en la precipitación y el aumento de la frecuencia de eventos de heladas estacionales.,
finalmente, nuestros resultados plantean preguntas importantes con respecto a la variación dentro de las especies, particularmente en relación con la predicción del impacto del cambio climático o la selección de las semillas a utilizar para la restauración. Las estrategias de germinación pueden variar dentro de las especies dependiendo de la población, el tiempo de floración, la masa de semilla, la inversión parental, el clima, la elevación y/o el ambiente materno y esta variación puede reflejar factores genéticos, epigenéticos y/o ambientales (Cochrane et al., 2015)., El aumento de la elevación explica la variación en el estado de germinabilidad y / o latencia de varias especies alpinas (Billings y Mooney, 1968; Dorne, 1981; Cavieres y Arroyo, 2000), pero véase (Hoyle et al., 2014). El estado de latencia reducida puede resultar de la terminación temprana del desarrollo de semillas como resultado de tensiones ambientales (Steadman et al., 2006; Hoyle et al., 2008b)., El examen de la variación en las estrategias de germinación dentro de las especies puede revelar el potencial de las especies para hacer frente y adaptarse a los cambios en el clima, por lo tanto, desempeñar un papel importante en la determinación de la supervivencia futura y la distribución de las especies y proporcionar información importante para la gestión y la conservación.
conclusiones
nuestro enfoque ha revelado la ocurrencia generalizada de latencia en la flora alpina australiana y demuestra que las estrategias de germinación varían dentro de una colección de semillas, así como entre colecciones., Además, hemos demostrado que si bien la estrategia de dormancia se conserva filogenéticamente y se correlaciona con la masa de semillas y la endosperma, es en gran medida independiente de los rasgos vegetativos y las características del área de distribución. Si bien no se puede extrapolar directamente a partir de nuestros resultados cómo el clima y los cambios asociados afectarán a la flora alpina, estos resultados pueden formar la base para el diseño de planes de propagación y la experimentación adicional sobre el impacto de los regímenes climáticos cambiantes., En particular, abogamos por una mayor investigación para comprender las causas de la variación dentro de las especies en la estrategia de germinación y el papel que esto puede desempeñar en la capacidad de las especies para resistir el cambio climático predicho.
accesibilidad de los datos
Los Datos Completos de germinación y la base de datos de rasgos se alojarán en DRYAD y en el Atlas of Living Australia.
Declaración de conflicto de intereses
los autores declaran que la investigación se realizó en ausencia de relaciones comerciales o financieras que pudieran ser interpretadas como un potencial conflicto de intereses.,
agradecimientos
reconocemos la financiación del Australian Research Council (Lp0991593) y el Jardín Botánico Nacional de Australia y sus amigos. Agradecemos a J. McAuliffe, S. Fethers, y S. Lawatsch de la ANBG. También agradecemos A M. Holloway-Phillips, M. Cardillo y S. Cunningham por sus aportes sobre las versiones anteriores del manuscrito y por su asesoramiento analítico, y a S. Venn por el acceso a los datos de rasgos.
Supplementary Material
Foley, M. E. (1992). Efecto de los azúcares solubles y el ácido giberélico en la ruptura de la latencia de embriones de avena salvaje (Avena fatua) extirpados., Weed Sci. 40, 208–214.
Google Scholar
Harden, G. J. (1990). Flora of New South Wales. Sydney, NSW: Royal Botanic Gardens.
Google Scholar
International Seed Testing Association. (2003). Hojas de trabajo sobre las pruebas de tetrazolio. Bassersdorf :STA.
Kullman, L. (2004). The changing face of the alpine world (en inglés). Glob. Change Newsl. 57, 12–14.
Google Scholar
Deja una respuesta