
El polimorfismo genético es la existencia de Estados alternativos del ADN, determinando la variación de los niveles más altos de integración del organismo. Hay diferentes tipos de modificaciones del genoma (mutaciones). Las más estudiadas son las sustituciones de nucleótidos en regiones codificantes y reguladoras.
Definición
El polimorfismo genético es la existencia, en una población, de varios estados alternativos de ADN, o alelos, en una posición definida en el genoma, o locusposición del gen en el cromosoma., En genética de poblaciones, un conjunto de genes homólogos (clase de homología). Dos genes son homólogos si coinciden con la meiosis. Plural: loci.. Esta definición tiene varios aspectos:
(1) Primero, el rasgo debe ser llevado por los cromosomas y ser transmisible.
(2) Entonces los alelos deben ser homólogos dos cromosomas o se dice que dos genes son homólogos si coinciden y se excluyen mutuamente de la meiosis. por su posición en el genoma, lo que excluye ciertos tipos de variación que se discutirán más adelante.,
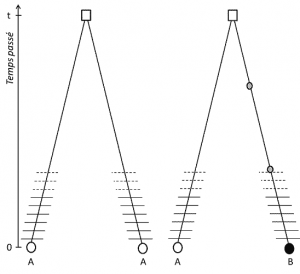
(3) pero dado que el rasgo es transmisible, la homología de posición también implica que los alelos son homólogos por descendencia; si son diferentes, es por lo tanto que una mutación (al menos) ha ocurrido en una de las líneas que los une a su último ancestro común (Figura 1).
(4) Finalmente, el polimorfismo genético se puede definir a la escala de la unidad más pequeña que compone el ADN: el sitio del nucleótido., Como resultado, cada variante de nucleótido puede determinar el polimorfismo de los niveles más altos de estructura biológica – el gen, la proteína y el fenotipo del individuo – que se convierten en escalas en las que se puede describir el polimorfismo genético (Figura 2).
tomemos el ejemplo del sistema sanguíneo ABO, que corresponde a motivos antigénicos parte del antígeno reconocido por un anticuerpo o receptor de linfocitos. También conocido como epítopo o determinante antigénico. El mismo antígeno puede tener varios epítopos (idénticos o diferentes) y así inducir una respuesta inmune variada., participa en la compatibilidad durante las transfusiones de sangre. Este es un caso de polimorfismo genético presente en poblaciones humanas, involucrando la proteína ABO glicosil transferaseenzima que permite la transferencia de residuos que contienen azúcares a proteínas. En el caso del sistema ABO, la glicosil transferasa A y B respectivamente inducen a un individuo a pertenecer al grupo A o B. cuando ambas glicosiltransferasas están presentes, el individuo es del grupo AB. que tiene tres alelos, A, B y O. Un individuo puede tener genotipos (AA), (ao); (BB), (BO); (oo) o (AB)., Su fenotipo será entonces,,;,; o, donde vemos que A y B «dominan» O (es decir, su expresión enmascara la de O), y que son co-dominantes entre sí (el heterocigoto entre A y B tiene una expresión reconocible,).
un polimorfismo se puede describir primero a la escala de la secuencia de ADN del locus que codifica una proteína. Algunos polimorfismos son» sinónimos», es decir, no cambian la secuencia de aminoácidos de la proteína; a menudo son los polimorfismos más numerosos. Los otros modifican los aminoácidos y se llaman polimorfismos de «reemplazo»., Los polimorfismos de reemplazo de ABO glicosil transferasa son de dos tipos: polimorfismos que cambian el aminoácido pero no la unidad antigénica (no interfieren con el fenotipo ABO de los individuos); y otros polimorfismos de reemplazo, que determinan el conjunto de fenotipos ABO de características observables de un individuo..
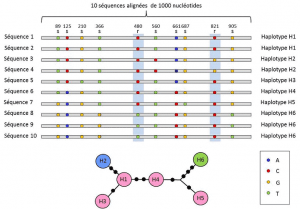
entre los niveles de nucleótidos y alelos, los análisis genéticos tienen en cuenta un nivel intermedio de descripción: la disposición lineal de los sitios variables en el locus. Estos arreglos, llamados haplotiposgrupo de alelos de diferentes loci ubicados en el mismo cromosoma Y generalmente se transmiten juntos. El haplotipo se forma por la contracción de la frase inglesa haploid genotype, o haploid genotype., Todos los genes ubicados en el mismo cromosoma y cuyos alelos se segregan durante la meiosis constituyen un haplotipo. Estos genes se denominan «genéticamente vinculados», son útiles en genética evolutiva porque permiten encontrar los vínculos genealógicos entre los alelos (Figura 2).
los polimorfismos de nucleótidos también existen en regiones intergénicas (fuera de las regiones codificantes). Algunos afectan las áreas donde la expresión génica está regulada y por lo tanto tienen expresión fenotípica. Otros no tienen ningún efecto conocido y se conocen como «silenciosos».,
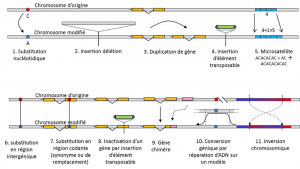
Además de las sustituciones de nucleótidos, el polimorfismo para un tipo particular de mutación, los microsatélites, se refiere a las variaciones en la longitud de un fragmento de ADN a menudo no codificante (Figura 3) debido a un cambio en el número de repeticiones en una breve secuencia repetida de nucleótidos, por ejemplo, CACACACA o TGTGTG.
no toda variación genética entra dentro de la definición de polimorfismo genético, ya que requiere la homología de posición de las alternativas (Figura 3)., Por lo tanto, hay secuencias repetidas obtenidas duplicando un gen, para las cuales dos repeticiones no son homólogas por posición. A menudo, tampoco podemos hablar de homología para elementos transponibles, porque generalmente cambian de posición durante las generaciones celulares, pueden multiplicarse e invadir el genoma. La homología de dos elementos transponibles no puede entonces definirse. Por otro lado, dos secuencias codificantes del mismo locus, una de las cuales está intacta, y la otra inactivada por la inserción de un elemento transponible, corresponden bien a una situación de homología., Un día, probablemente consideraremos hablar de polimorfismo para las modificaciones de los llamados cromosomas «epigenéticos», que a veces se transmiten a través de varias generaciones celulares, células somáticas calificadas no reproductivas, o soma. Una mutación somática, que afecta a un gen de una célula somática, desaparece con el individuo portador. o germinalacalifica los gametos. Una mutación de la línea germinal puede transmitirse a su descendencia., como la metilación. Queda por evaluar su importancia en la evolución de las poblaciones., La limitación de la noción de polimorfismo genético a ciertas categorías de variación – esencialmente sustituciones de nucleótidos – proviene de la gran utilidad de esta última en la exploración de la historia de las poblaciones naturales. Se prestan al modelado matemático de la evolución.
la palabra polimorfismo significa «varias formas». Se opone al monomorfismo, que indica la ausencia de variación. En el vocabulario de la biología, esta oposición se ha utilizado durante mucho tiempo, antes que la genética, para señalar la coexistencia de varios tipos de individuos en la misma especie, por ejemplo., caste polymorphism (queen and workers) in social insects; seasonal polymorphism (fur changes) in some Arctic mammals. Estos casos no caen bajo el polimorfismo genético, un concepto más reciente con un significado más estrecho y preciso. Las variaciones continuas (por ejemplo, la variación de tamaño) tampoco entran en la categoría de polimorfismo, ya que no presentan alternativas distintas. Sin embargo, los locus que afectan el tamaño caen dentro de esta definición., A continuación discutiremos las variaciones genéticas más estudiadas porque son las más importantes para la evolución, las sustituciones de nucleótidos, y luego discutiremos la variación de fenotipos.
Medición
no es suficiente decir que un lugar es más o menos variable, sin dar una estimación cuantitativa a este juicio. Un investigador puede estudiar la variación a diferentes escalas. Si solo está interesado en los alelos de una proteína, medirá la diversidad alélica, simbolizada por ‘H’. Si está interesado en la diversidad del ADN, medirá la diversidad de nucleótidos, simbolizada por ‘π’.,
la diversidad de alelos H se define como la probabilidad de dibujar dos alelos diferentes durante dos muestreos con replacementPerforming un sorteo sucesivo con entrega de tokens p en una urna que contiene tokens n, significa tomar un primer token, leer su valor, ponerlo de nuevo en la urna, tomar un segundo token, leer su valor, ponerlo de nuevo en la urna, etc., hasta el token pth. Esto significa elegir objetos p entre n con repetición (Puede elegir el mismo objeto varias veces) y en orden (el orden en el que elija los objetos es importante)., El número de sorteos sucesivos con fichas entre n es: n × n × n × × × n = np.. Si llamamos pi La frecuencia del alelo de rango i, mostramos que la probabilidad de elegir dos veces el mismo alelo es F = Σ pi2. La diversidad alélica de la muestra es entonces su complemento a 1, es decir:
H = 1 – Σ pi2 (1)
esto se aplica tanto a los alelos proteicos como a los haplotipos. Esto se llama diversidad haplotípica. También se puede llamar heterocigosidad, porque en el caso de un locus diploide, da la frecuencia esperada de heterocigotos.,
la diversidad de nucleótidos π es el equivalente de H A la escala de cada nucleótido. Se calcula como el promedio del número de diferencias de nucleótidos entre secuencias en una muestra tomada de dos en dos (δij), dividido por la longitud del fragmento de ADN en el número de nucleótidos (L).
π = average (δij) /l (2) (ver referencia)
Este valor difiere entre especies. En las partes codificantes del genoma humano, dos cromosomas seleccionados aleatoriamente de la población difieren en promedio en un nucleótido por mil., En la mosca de la fruta (Drosophila melanogaster) esta diferencia es de aproximadamente una en cien. La mosca es diez veces más variable que el hombre. Cuando estos valores están relacionados con el tamaño de las regiones codificantes (alrededor de 15.500 genes en Drosophila, al menos 22.000 en humanos), o incluso genomas (140 millones de pares de bases por genoma haploide en moscas, alrededor de 3.200 millones o veinte veces más en humanos), el número de sitios polimórficos es astronómico y resulta en que nadie nacido de la generación sexualizada en ninguna especie es, ha sido y será genéticamente idéntico a otro pasado, presente o futuro., El poder de información del polimorfismo del ADN es inmenso. En medicina forense, los detectives pueden identificar a cualquier sospechoso de dieciséis locuses de microsatélites.
historia
el término variación está en el título de los dos primeros capítulos del libro de Darwin El Origen de las especies (1859). Darwin introdujo esta noción en las ciencias naturales en un momento en que las leyes de la herencia biológica seguían siendo un misterio, y le dedicó otro libro importante, Variation in Animals and Plants (1868)., Convencido de que la evolución se trataba de pequeñas variaciones que hacían poca diferencia en la adaptación de los organismos a sus condiciones de vida, atribuyó una importancia crucial a las pequeñas variaciones cuantitativas, lo que llevó a sus sucesores (especialmente Karl Pearson) a fundar la biometría de la medición de la vida. Se refiere en un sentido muy amplio al estudio cuantitativo de los seres vivos.. Pero el redescubrimiento de las leyes de Mendel en 1900 cambió el interés a las variaciones discontinuas.
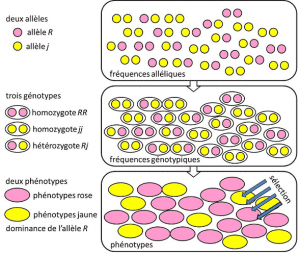
de 1908 a 1930, la genética de poblaciones fue esencialmente una disciplina teórica que buscaba reconciliar el darwinismo con la herencia mendeliana basada en la transmisión de un solo gen bajo un modo dominante, recesivo o ligado a cromosomas sexuales X (O y). Se refiere a un rasgo genético con determinismo simple, por una pareja o un pequeño número de parejas de genes., y donde las probabilidades jugaron un papel importante. La genética es contra-intuitiva., Predice que no hay reproducción del Padre en su descendiente, porque el genotipo de este último resulta de la Lotería de la separación de los alelos antes de la fusión de los dos semigenomas parentales. Se dio cuenta de que estas son las frecuencias alélicasfrecuencia en la que el alelo de una variante se encuentra en una población. Expresado como proporción o porcentaje. La suma de las frecuencias alélicas de todos los alelos de un gen en una población es, por definición, igual a 1. En genética de poblaciones, las frecuencias alélicas representan la diversidad genética a nivel de población o especie., que se transmiten entre generaciones, no genotipos o fenotipos. Estas frecuencias son más o menos estables de una generación a la siguiente y producen, más allá del enredo múltiple de filiaciones entre individuos, las mismas frecuencias genotípicas estructura genética de la población. Se determina a partir de las frecuencias alélicas (Figura 4). La distribución de los genotipos poblacionales, asociada a la varianza, es por lo tanto el único factor predecible., Alrededor de 1930, los tres teóricos Ronald Fisher, JBS Haldane y Sewall Wright ayudaron a crear la noción de la población mendeliana cuya herencia obedecía las leyes de Mendel. . En este contexto, la evolución necesariamente asocia tres niveles estructurales: el gen, el individuo y la población. Esta asociación de tres vías se puede resumir con la siguiente fórmula: la evolución de la población es un cambio en la frecuencia de los alelos (escala génica, pero también escala de grupo), un cambio que depende de la clasificación de fenotipos por selección (escala individual)., La investigación empírica de genética de poblaciones se desarrolló entonces, pero estuvo limitada durante mucho tiempo por nuestra ignorancia de cómo funcionan los cromosomas (la estructura del ADN se entendió en 1953 y su secuenciación comenzó en 1977), y tuvo que recurrir al estudio de los pocos polimorfismos visibles, como la coloración del elytra de la mariquita o la concha de los caracoles (Véase Focus The great snail debate). Hubo mucho debate entre los genetistas sobre si las poblaciones naturales eran generalmente polimórficas o monomorfas, y si el polimorfismo era en sí mismo beneficioso o no., El estudio de la variación de la escala macromolecular tuvo que esperar hasta 1966 (para las proteínas) y 1983 (para el ADN) antes de responder a estas preguntas.
variaciones cuantitativas y heredabilidad
Cuando miramos a nuestro alrededor, vemos diferencias físicas entre los seres, algunas complejas como rasgos faciales, otras fáciles de medir como peso o altura. El sentido común sugiere que son parcialmente transmisibles, aunque de una manera que es difícil de especificar. Estos rasgos son a menudo polígenicunder la dependencia de muchos genes. Estamos hablando de herencia poligénica. La Diabetes es una enfermedad poligénica.,, lo que significa que están influenciados por muchos locuses. La genética del desarrollo, y su aplicación a la comparación de especies (evo-devo, por «evolución del desarrollo») desentrañan la red de interacciones complejas que hacen que durante el desarrollo las células del cuerpo, todas con idéntico bagaje genético, se diferencien por epigenesis para dar diferentes tejidos. Todavía no está claro cómo se construyen los personajes complejos. Los fenotipos medibles se pueden estudiar utilizando la biometría, basada en las propiedades de las distribuciones estadísticas., Un valor con propiedades útiles es la varianciaen una muestra de individuos en los que se midió un rasgo dado, la varianza es la diferencia entre la media del cuadrado de valores y el cuadrado de la media de los valores. Esta medida, siempre positiva, indica la dispersión de los individuos. . Cuando varias causas independientes determinan la variación de una línea, sus varianzas son aditivas y su suma da la varianza de la línea. Si no son independientes, la suma de las covarianzas se suma a la suma de las varianzas., La varianza total Vt de un rasgo fenotípico es tal que :
Vt = Vga+Vgd+Vgd+VGI+Ve
VGA, varianza genética aditiva, es la suma de las varianzas debidas a cada locus independientemente de los otros; Vgd, varianza genética dominante, resulta de la interacción de los dos alelos del mismo locus; Vgi, varianza de interacción entre locus, resulta del hecho que los locuses del mismo individuo interactúan; ve, varianza del ambiente, se asume aquí independiente de varianza genética.,
la varianza genética de dominancia es común a dos hijos de los mismos padres, porque comparten los efectos de dominancia cuando en un locus dado han recibido los mismos alelos de ambos padres, lo que les sucede en uno de cada cuatro locus. No comparten estos efectos con sus padres, por lo que son más «similares» entre sí de lo que son con sus padres, a pesar del hecho de que comparten tantos genes entre sí. Ciertamente, un padre puede, por casualidad, haber recibido los mismos dos alelos que uno de sus hijos en un lugar determinado., Esta posibilidad depende de la frecuencia de los alelos en la población. Vemos que el grado de similitud entre un niño y su padre para un rasgo dado es una propiedad pública.
Más generalmente, todos los componentes de la varianza genética (Vga+Vgd+Vgi) pueden variar entre poblaciones. En las líneas de reproducción, un caso común es la endogamia, que agota la varianza genética: como resultado, un rasgo puede tener el mismo promedio en dos poblaciones, pero tener una variación genética diferente. La capacidad de un criador para seleccionar un rasgo es, por lo tanto, una característica de la línea, no del rasgo.,
la relación entre padres e hijos se mide por heredabilidad h2. Se define como la proporción de la varianza genética aditiva a la varianza total :
h2 = Vga/Vt

(2) correlación de 0, donde el tamaño de los hijos es independiente del de los padres. Un padre medio valioso ha producido un hijo de valor indiferente, dentro de la distribución de los hijos. Las coordenadas de los puntos se encuentran en un círculo; la expectativa de su valor se encuentra en una línea paralela al eje de los padres (d)., (3) correlación intermedia padre-hijo entre 0 y 1: Las coordenadas de los puntos se encuentran en una elipse. La expectativa de su valor ya no se encuentra en la bisectriz de los ejes de coordenadas (que también es el eje mayor de la elipse de distribución), sino en la línea de regresión r. un padre promedio de valor ha producido un hijo de valor b (entre 0 y a) con una cierta varianza. Se observa que el caso (3) es intermedio entre los casos (1) y (2)
se muestra bajo varios supuestos en la Figura 5., Heredabilidad es la capacidad de determinar si una característica puede ser seleccionada. La figura 5-3 muestra que un criador podrá, dentro de una generación cruzada, mover el promedio de un personaje del valor 0 al valor b Si selecciona individuos con valor a. podemos demostrar que b = h2a. para seleccionar un rasgo, es útil aumentar la relación de heredabilidad estandarizando el entorno (reduciendo así la varianza ambiental), y colocándose en las condiciones ambientales que favorecen la aparición del rasgo., El genetista Waddington ha demostrado que, en la naturaleza, un cambio de entorno puede revelar nuevos rasgos que luego serán seleccionados durante la evolución.
referencias y notas
imagen de portada. Conchas de caracol Cepaea nemoralis & Cepaea hortensis. André Künzelmann, UFZ.,
esta fórmula se puede escribir de una manera más convencional como: \pi =\frac{n(n-1))}{2L}\sum _{i=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
siendo una muestra de individuos en los que se midió el rasgo estudiado, la varianza se puede definir como la diferencia entre la media del cuadrado de valores y el cuadrado de la media de los valores. Esta medida, siempre positiva, indica la dispersión de los individuos.
esto es heredabilidad en el sentido estricto del término. Heredabilidad en un sentido amplio es la relación entre la suma de varianzas genéticas y varianza total., Esta fórmula da heredabilidad con ambos padres( el «padre promedio»); la estimación con un solo padre daría h2/2.
The Environmental Encyclopedia of the Environment by the Association des Encyclopédies de l’environnement et de l’énergie (www.a3e.fr), vinculado contractualmente a la Universidad de Grenoble Alpes y Grenoble INP, y patrocinado por la Academia francesa de Ciencias.
para citar este artículo:VEUILLE Michel (2021), genetic polymorphism and variation, Encyclopedia of the Environment, url : https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
los artículos de la Enciclopedia del medio ambiente están disponibles bajo los Términos de la Licencia Creative Commons BY-NC-SA, que autoriza la reproducción sujeta a: citar la fuente, no hacer uso comercial de ellos, compartir idénticas condiciones iniciales, reproducir en cada reutilización o distribución la mención de esta Licencia Creative Commons BY-NC-SA.
Deja una respuesta