DNA: n ja RNA
kaksi päätyyppiä nukleiinihapot ovat deoksiribonukleiinihapon (DNA) ja ribonukleiinihappo (RNA). DNA on kaikissa elävissä eliöissä esiintyvä geneettinen materiaali, joka vaihtelee yksisoluisista bakteereista monisoluisiin nisäkkäisiin. DNA: ta esiintyy eukaryoottien tumassa sekä organelleissa, kloroplasteissa ja mitokondrioissa. Prokaryooteissa DNA: ta ei suljeta kalvomaiseen kuoreen.
solun koko geneettinen sisältö tunnetaan sen genomina, ja genomitutkimus on genomitutkimusta., Eukaryoottisissa soluissa mutta ei prokaryooteissa DNA muodostaa histoniproteiinien kanssa kompleksin, josta muodostuu eukaryoottisten kromosomien aine kromatiini. Kromosomissa voi olla kymmeniätuhansia geenejä. Monet geenit sisältävät tietoa proteiinituotteiden valmistamiseksi; muut geenit koodaavat RNA-tuotteita. DNA ohjaa kaikkea solutoimintaa kääntämällä geenit ”päälle” tai ” pois päältä.”
muu nukleiinihappo, RNA, osallistuu enimmäkseen proteiinisynteesiin. DNA-molekyylit eivät koskaan poistu tumasta, vaan käyttävät välittäjää kommunikoidakseen muun solun kanssa., Tämä välittäjä on lähetti-RNA (mRNA). Muut RNA—tyyppiset rRNA—, tRNA-ja mikroRNA-lajit osallistuvat proteiinisynteesiin ja sen säätelyyn.
DNA ja RNA koostuvat nukleotideina tunnetuista monomeereista. Nukleotidit yhdistyvät toisiinsa muodostaen polynukleotidin, DNA: n tai RNA: n. Kunkin nukleotidin koostuu kolmesta osasta: typpipitoisen emäksen, pentoosi (viisi-carbon) sokeri ja fosfaatti-ryhmä (Luku \(\PageIndex{1}\)). Jokainen nukleotidin typpiemäs kiinnittyy sokerimolekyyliin, joka kiinnittyy yhteen tai useampaan fosfaattiryhmään.,
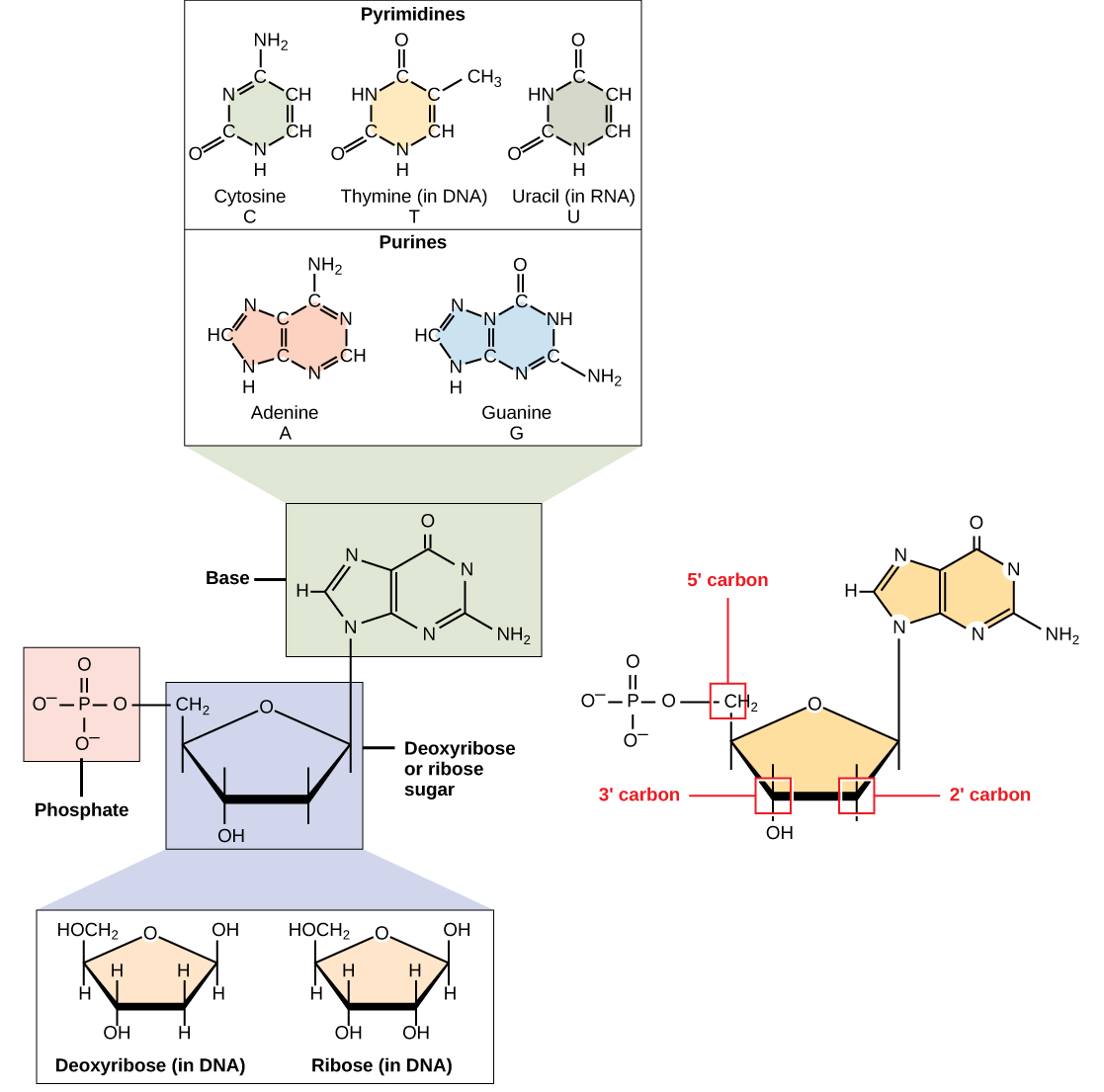
typpipitoiset emäkset, tärkeitä osia nukleotidit, ovat orgaanisia molekyylejä, ja ne ovat niin nimeksi, koska ne sisältävät hiiltä ja typpeä. Ne ovat emäksiä, koska ne sisältävät amino-ryhmä, joka on mahdollista sitovat ylimääräistä vetyä, ja siten vähentää vetyionipitoisuus sen ympäristössä, jolloin se on enemmän perus. Kunkin nukleotidin DNA sisältää yhden neljästä mahdollisesta typpipitoiset emäkset: adeniini (A), guaniini (G), sytosiini (C) ja tymiini (T).
adeniini ja guaniini luokitellaan puriineiksi., Puriinin perusrakenne on kaksi hiilityppirengasta. Sytosiini, tymiini ja urasiili ovat luokiteltu pyrimidines, joka on hiili-typpi-rengas, koska niiden ensisijainen rakenne (Luku \(\PageIndex{1}\)). Jokaisessa näistä hiilen ja typen perusrenkaista on siihen kiinnittyneet erilaiset funktionaaliset ryhmät. Molekyylibiologian pika -, ja typpipitoiset emäkset ovat yksinkertaisesti tunnetaan niiden symbolit A, T, G, C, ja U. DNA sisältää A, T, G ja C, kun taas RNA sisältää A, U, G ja C.
pentoosisokeri DNA on deoxyribose, ja RNA, sokeri on riboosi (Luku \(\PageIndex{1}\))., Ero sokerit on läsnäolo hydroksyyli-ryhmän toinen hiili riboosi-ja vety toisen hiilen deoxyribose (niin deoxyribose on ”puuttuu” yksi -OH-ryhmä). Hiiliatomien sokerin molekyyli on numeroitu kuten 1′, 2′, 3′, 4′, ja 5′ (1′ luetaan ”yksi prime”). Fosfaatti jäännös on kiinnitetty hydroksyyli-ryhmä 5′ hiili yksi sokeria ja hydroksyyliryhmä 3′ hiili sokeria seuraavan nukleotidin, joka muodostaa 5′-3′ phosphodiester sidos., Se phosphodiester sidos ei ole muodostettu yksinkertainen nestehukka reaktio kuin muut kytkökset yhdistävät monomeerit vuonna makromolekyylit: sen muodostumista liittyy poistaminen kaksi fosfaattiryhmää. Polynukleotidilla voi olla tuhansia tällaisia fosfodiesterisidoksia.
Chargaff: n Säännöt
Kun Watson ja Crick esitetty 1940-luvun määrittää DNA: n rakenteen, se oli jo tiedossa, että DNA on tehnyt sarjassa neljä erilaista molekyylejä, nimeltään emäkset tai nukleotidien: adeniini (A), tymiini (C), tymiini (T), guaniini (G)., Watson ja Crick myös tiesi Chargaff: n Sääntöjä, jotka olivat joukon huomautuksia, suhteellinen määrä kunkin nukleotidin, joka oli läsnä lähes kaikki ote DNA: ta. Chargaff oli havaittu, että kunkin lajin runsaus oli sama kuin T, ja G on sama kuin C. Tämä oli välttämätöntä Watson & Crick malli.
Esimerkiksi \(\PageIndex{1}\)
Chargaff määräytyy koostumus nukleiinihapot näytteitä eri lajeja, kuten prokaryooteissa ja eukaryooteissa. Yhdessä bakteerinäytteessä adeniinin osuus oli 15.,5% (TIEDOT mukautettu Vischer et al, 1949). Mikä osuus guaniinia olisi ollut tässä otoksessa ja miksi?
liuos
koska T-pareilla T: n määrän tulee olla suurin piirtein A eli noin 15,5%. Näin Ollen A + T = 15,5 + 15,5 = 31%.
prosenttiluku G + C = 100% – 31% = 69%. Koska G paria C, määrä kunkin näistä pitäisi olla suurin piirtein yhtä suuri, joten noin 34,5% kukin.,
DNA-Double-Helix-Rakenne
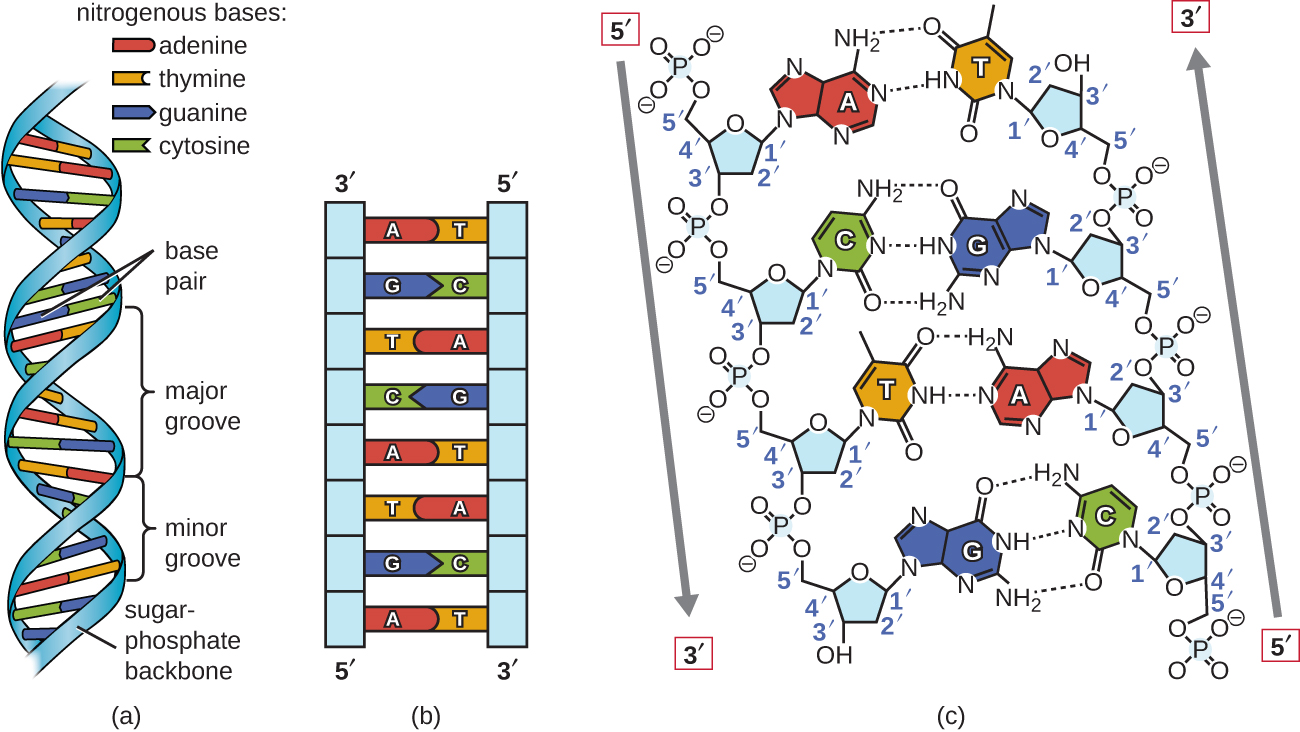
Käyttämällä suhteellisia metalli malleja yksittäisten nukleotidien, Watson ja Crick päätellä, rakenne, DNA: n, joka oli sopusoinnussa Chargaff: n Sääntöjä ja x-ray crystallography, että tiedot on saatu (joitakin kiista) toinen tutkija nimeltä Rosalind Franklin. Watson ja Crick on kuuluisa kaksoiskierre, kumpikin osa sisältää DNA: n emäkset yhdistetty kovalenttisten joukkovelkakirjojen sokeri-fosfaatti selkäranka., Koska yksi puolella jokainen sokerimolekyyli on aina kytketty vastakkaisella puolella ensi sokeria molekyyli, jokainen DNA: ta on napaisuus: nämä ovat nimeltään 5′ (5-prime) pää-ja 3′ (3-prime) end mukaisesti nimikkeistön hiilivetyjä vuonna sokerit. Nämä kaksi osa-double helix ajaa anti-rinnakkain (eli vastakkaiseen) suuntaan, jossa 5′ loppuun yhden lohkon vieressä 3′ päähän toisen lohkon. Kaksoiskierre on oikeakätinen kierre, (pikemminkin kuin vasenkätinen kierre, joka on usein edustettuna väärin suosittu media)., DNA: n emäkset ulottuvat selkäranka kohti keskustaa helix, pari emäksiä kunkin lohkon muodostavat vetysidoksia, jotka auttavat pitämään kaksi säiettä yhteen. Useimmissa olosuhteissa, kaksi säikeet ovat hieman offset, joka luo merkittävän uran yksi kasvot double helix, ja pieni ura muita. Koska rakenteen perusteet, voi vain muodostaa vetysidoksia T ja G voi vain muodostaa vetysidoksia C (muista Chargaff: n Säännöt)., Kunkin juosteen sanotaan siis täydentävän toisiaan, joten jokainen juoste sisältää myös riittävästi tietoa, jotta se toimisi mallina toisen synteesille. Tämä täydentävä irtisanominen on tärkeää DNA: n replikoinnissa ja korjaamisessa.
vain tietyntyyppiset emäsparit ovat sallittuja. Esimerkiksi tietty puriini voi pariutua vain tietyn pyrimidiinin kanssa. Tämä tarkoittaa tölkkiparia T: n kanssa, ja G voi parittaa C: n kanssa, kuten kuvassa \(\PageIndex{2}\) esitetään. Tätä kutsutaan base complementary ruleksi. Toisin sanoen DNA-säikeet täydentävät toisiaan., Jos yhden lohkon jono on AATGGCC, täydentävällä lohkolla olisi sekvenssi TTAACCG. DNA: n replikaation aikana jokainen juoste kopioidaan, jolloin tytär DNA: n kaksoiskierre sisältää yhden vanhemman DNA-juosteen ja vasta syntetisoidun juosteen.
Thinking eteenpäin harjoitus \(\PageIndex{1}\)
A-mutaatio tapahtuu, ja sytosiini korvataan adeniini. Miten tämä vaikuttaa DNA: n rakenteeseen?,
Vastaus
Adeniini on suurempi kuin sytosiini ja ei voi emäsparin oikein guaniini vastapuolen strand. Tämä saa DNA: n pullistumaan. DNA-korjausentsyymit saattavat tunnistaa pullistuman ja korvata virheellisen nukleotidin.
Vastaa