
Geneettinen polymorfismi on olemassa vaihtoehtoisia todetaan, DNA, määritetään vaihtelu organismi on korkeampi integraation. On olemassa erilaisia genomin muutoksia (mutaatioita). Tutkituimpia ovat nukleotidisubstituutiot koodaus-ja sääntelyalueilla.
Määrittely
Geneettinen polymorfismi on olemassa, väestön, useita vaihtoehtoisia todetaan, DNA, tai alleeleja, klo määritelty asema genomin, tai locusPosition geeni kromosomissa., Populaatiogenetiikassa joukko homologisia geenejä (homologian Luokka). Kaksi geeniä on homologisia, jos ne sopivat meioosiin. Monikko: loci.. Tämä määritelmä on useita näkökohtia:
(1) Ensin, sen piirre on tehtävä, jonka kromosomit ja olla tarttuvia.
(2) Sitten alleeleja on homologousTwo kromosomit tai kaksi geenien sanotaan olevan homologisia, jos ne sopivat ja toisiaan sulje pois toisiaan meioosin. heidän asemaansa genomissa, mikä sulkee pois tietyntyyppiset variaatiot, joista keskustellaan myöhemmin.,
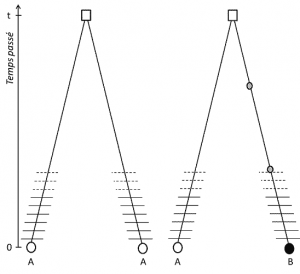
(3), Mutta koska ominaisuus on siirrettävissä, kanta homologia tarkoittaa myös sitä, että alleelit ovat homologisia, jonka laskeutuminen; jos ne ovat erilaisia, se on siksi, että mutaatio (ainakin) on tapahtunut yhden viivan, joka yhdistää heidät heidän viimeinen yhteinen esi-isä (Kuva 1).
(4) Lopuksi, geneettinen polymorfismi voidaan määritellä mittakaavassa pienin yksikkö säveltäminen DNA: nukleotidin sivuston., Tämän seurauksena jokainen nukleotidi-muunnos voidaan määrittää polymorfismi korkeampi biologinen rakenne – yksilö on geeni -, proteiini-ja fenotyyppi – jotka tulevat asteikot, jossa geneettinen polymorfismi voidaan kuvata (Kuva 2).
Otetaan esimerkiksi ABO veren järjestelmä, joka vastaa antigeeninen motivesPart antigeenin tunnustettu vasta-aine tai lymfosyyttien reseptorin. Tunnetaan myös epitooppina tai antigeenisenä determinanttina. Samalla antigeenillä voi olla useita epitooppeja (identtisiä tai erilaisia) ja siten aiheuttaa vaihtelevan immuunivasteen., osallistuu yhteensopivuuteen verensiirtojen aikana. Kyseessä on geneettinen polymorfismi läsnä väestöryhmiin, joissa proteiini ABO glykosyyli transferaseEnzyme jolloin siirto jätteet, joissa sokerit ja proteiinit. ABO-järjestelmässä glykosyylitransferaasi A ja B indusoivat yksilön kuulumaan A-tai B-ryhmään. jolla on kolme alleelia, A, B ja O. yksilöllä voi olla (AA), (AO), (BB), (BO), (OO) tai (AB) genotyyppejä., Sen fenotyyppi on sitten ,,; ,;, tai jos näemme, että A ja B ”hallitsevat” O (eli niiden ilmaisun naamiot, jotka O), ja että he ovat yhteistyössä hallitseva keskenään (heterotsygoottisessa A ja B välillä on tunnistettava ilme, ).
polymorfismi voidaan kuvata ensin proteiinia koodaavan lokuksen DNA-sekvenssin asteikolla. Jotkut polymorfismit ovat ”synonyymi”, eli ne eivät muuta aminohappo sekvenssin proteiinia; he ovat usein eniten polymorfismeja. Muut muokkaavat aminohappoja ja niitä kutsutaan ”korvaaviksi” polymorfismeiksi., Vaihto polymorfismeja ABO glykosyyli transferaasin ovat kahdenlaisia: polymorfismeja, joka muuttaa aminohapon mutta ei antigeeni-yksikkö (ne eivät häiritse ABO yksilöiden fenotyyppi); ja muut vaihto polymorfismeja, jotka määrittävät ABO phenotypeSet havaittavissa ominaisuudet yksilön..
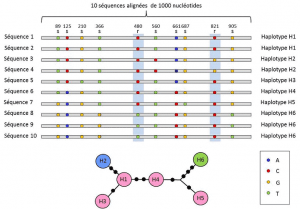
Välillä nukleotidi-ja-alleeli tasoilla, geneettiset analyysit ottaa huomioon keskitason kuvaus: lineaarinen järjestely muuttuja sivustoja locus. Nämä järjestelyt, joita kutsutaan haplotyyppiryhmäksi eri lokusten alleeleja, jotka sijaitsevat samassa kromosomissa ja siirtyvät yleensä yhdessä. Haplotyyppi on englantilaisen fraasin haploidigenotyypin eli haploidigenotyypin supistuminen., Kaikki geenit sijaitsevat samassa kromosomissa ja joiden alleelien erillään yhdessä aikana meioosin muodostavat haplotyyppi. Nämä geenit ovat nimeltään ”periytyvä”, ovat hyödyllisiä in evolutionary genetics, koska niiden avulla löytää sukututkimuksen yhteyksiä alleeleja (Kuva 2).
Nukleotidipolymorfismeja on myös intergeenisillä alueilla (koodausalueiden ulkopuolella). Jotkut vaikuttavat alueisiin, joilla geenien ilmentymistä säännellään ja joilla on siksi fenotyyppinen ilmentymä. Muilla ei ole tunnettua vaikutusta, ja niistä käytetään nimitystä ”Hiljainen”.,
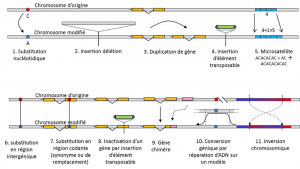
lisäksi nucleotide vaihdot, polymorfismi tietyn tyyppinen mutaatio, mikrosatelliitit, viittaa vaihtelut pituus on usein ei-koodaava DNA-fragmentti (Kuva 3), koska muutos määrä toistoja lyhyessä toistetaan järjestyksessä nukleotidien, esim. CACACACA, tai TGTGTG.
Ei kaikki geneettinen vaihtelu määritelmään geneettinen polymorfismi, koska se vaatii kanta homology vaihtoehtoja (Kuva 3)., Siten on olemassa toistuvia sekvenssejä, jotka saadaan monistamalla geeni, jolle kaksi toistoa eivät ole homologisia per asema. Usein, emme voi puhua homology varten transposonien joko, koska ne yleensä vaihtaa asentoa aikana solu sukupolvien, voi lisääntyä ja hyökätä genomin. Tämän jälkeen ei voida määritellä kahden transposoituvan alkuaineen homologiaa. Toisaalta, kaksi koodausta sekvenssit saman lokuksen, joista yksi on ehjä, ja muut inaktivoitu lisäämällä olentojen elementti, vastaavat hyvin homology tilanne., Eräänä päivänä, me luultavasti harkita puhumme polymorfismi muutoksia niin sanottu ”epigeneettiset” – kromosomia, jotka ovat joskus lähetetään useita solu sukupolvien, somaticQualifies ei-sukusolujen tai soma. Somaattisen solun geeniin vaikuttava somaattinen mutaatio katoaa kantajayksilön mukana. tai sukusoluja. Sukusolumutaatio voi tarttua sen jälkeläisiin., kuten metylaatio. Niiden merkitystä populaatioiden kehityksessä on vielä arvioitava., Rajoitus käsite geneettinen polymorfismi tietyt luokat vaihtelu – lähinnä nukleotidin vaihdot – tulee suuri apuohjelma, jälkimmäinen tutkia historiaa luonnon populaatiot. Ne soveltuvatkin evoluution matemaattiseen mallintamiseen.
sana polymorfismi tarkoittaa ”useita muotoja”. Se vastustaa monomorfismia, mikä osoittaa variaation puuttumisen. Sanasto biologian, tämä oppositio on jo pitkään käytetty, ennen kuin genetiikka, signaalin rinnakkaiselo useita erilaisia yksilöiden saman lajin, esim., kastin polymorfismi (kuningatar ja työläiset) sosiaalisissa hyönteisissä; kausiluonteinen polymorfismi (turkismuutokset) joillakin arktisilla nisäkkäillä. Nämä tapaukset eivät kuulu geneettinen polymorfismi, uudempi käsite on suppeampi ja tarkempi merkitys. Jatkuvat vaihtelut (esim.kokovaihtelu) eivät myöskään kuulu polymorfismin luokkaan, koska niissä ei ole erillisiä vaihtoehtoja. Kokoon vaikuttavat lokukset kuuluvat kuitenkin tämän määritelmän piiriin., Aiomme keskustella alla eniten tutkittu geneettistä vaihtelua, koska ne ovat tärkeimmät kehitys -, nukleotidi-vaihdot, ja sitten me keskustelemme vaihtelu fenotyyppeihin.
mittaus
ei riitä, että todetaan lokuksen muuttuvan enemmän tai vähemmän antamatta tälle tuomiolle määrällistä arviota. Tutkija voi tutkia vaihtelua eri asteikoilla. Jos hän on kiinnostunut vain alleelia proteiini, hän tulee mitata alleeliset monimuotoisuus, symboloi ”H”. Jos hän on kiinnostunut DNA: n monimuotoisuudesta, hän mittaa nukleotidien monimuotoisuutta, jota symboloi ”π”.,
– alleeli monimuotoisuus H on määritelty todennäköisyys piirustus kaksi eri alleelia aikana kaksi näytteenotto replacementPerforming peräkkäiset piirtää toimitus p kuponkia uurnassa, joka sisältää n kuponkia, tarkoittaa, että otetaan ensimmäinen token, lukeminen sen arvo, laitat sen takaisin uurnassa, kun toinen token, lukeminen sen arvo, laitat sen takaisin uurnassa, jne., kunnes pth token. Tämä tarkoittaa P-objektien valintaa n: n joukossa toistolla (voit valita saman objektin useita kertoja) ja järjestyksessä (järjestys, jossa valitset objektit on tärkeä)., Niiden peräkkäisten arvontojen lukumäärä, joissa on poletteja n: n joukossa, on: n × n × n × … × n = np.. Jos kutsumme pi taajuus alleeli rank i, osoitamme, että todennäköisyys poiminta kaksi kertaa sama alleeli on F = Σ pi2. Se alleeliset monimuotoisuus näyte on sitten sen täydennys 1, eli:
S = 1 – Σ pi2 (1)
Tämä koskee sekä proteiinia alleelit ja haplotyypit. Tätä kutsutaan haplotyyppiseksi monimuotoisuudeksi. Sitä voidaan kutsua myös heterotsygositeetiksi, koska diploidisen lokuksen tapauksessa se antaa heterotsygoottien odotetun taajuuden.,
nukleotidin monimuotoisuus π vastaa S mittakaavassa kunkin nukleotidin. Se lasketaan keskimääräinen lukumäärä nukleotidin eroja sekvenssit näyte otetaan kaksi kaksi (δij), jaettuna pituus DNA-fragmentti osaksi nukleotidien lukumäärä (L).
π = keskiarvo (δij)/L (2) (ks. viite )
Tämä arvo vaihtelee lajien välillä. Ihmisen genomin koodausosissa populaatiosta satunnaisesti valitut kaksi kromosomia eroavat keskimäärin yhdellä nukleotidilla tuhatta kohden., Hedelmäkärpäsessä (Drosophila melanogaster) ero on noin yksi sadasta. Kärpänen on siis kymmenen kertaa ihmistä vaihtelevampi. Kun nämä arvot liittyvät koko koodaus alueilla (noin 15,500 geenejä Drosophila, vähintään 22 000 ihmisellä), tai jopa genomit (140 miljoonaa emäsparia kohti haploidi genomin lentää, noin 3200 euroa tai kaksikymmentä kertaa enemmän ihmisiä), määrä polymorfinen sivustoja on tähtitieteellinen ja tuloksia ei ole yksi on syntynyt seksualisoitu sukupolvi tahansa laji on, on ollut ja tulee olemaan geneettisesti identtinen toisen menneisyyttä, nykyisyyttä tai tulevaisuutta., DNA-polymorfismin informaatiovoima on valtava. Tutkijat voivat tunnistaa epäillyn kuudestatoista mikrosatelliittipaikasta.
Historia
termi vaihtelu on otsikko, että kaksi ensimmäistä lukua Darwinin kirjan Lajien synty (1859). Darwin toi tämän käsityksen luonnontieteisiin aikana, jolloin biologisen perinnöllisyyden lait jäivät mysteeriksi ja omisti sille toisen tärkeän kirjan, eläinten ja kasvien vaihtelun (1868)., Vakuuttunut siitä, että kehitys oli pieniä eroja, että juurikaan eroa mukauttaminen organismien niiden elinoloja, hän kiinnitetty ratkaisevan tärkeitä pieniä määrällisiä eroja, mikä sai hänen seuraajansa (erityisesti Karl Pearson) löytyi biometryScience elämän arvostaminen. Viittaa hyvin laajassa merkityksessä elävien olentojen kvantitatiiviseen tutkimukseen.. Mutta Mendelin lakien uudistaminen vuonna 1900 siirsi kiinnostuksen epäjatkuviin muunnelmiin.
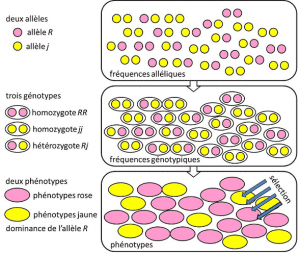
Vuodesta 1908 vuoteen 1930, populaatiogenetiikka oli lähinnä teoreettinen kurinalaisuutta pyrkiä sovittamaan yhteen Darwinismin kanssa Mendelin heredityHeredity perustuu lähetyksen yksittäinen geeni on dominoiva, resessiivinen tai sukupuoli kromosomi-linked-tila X (tai Y). Viittaa geneettiseen ominaisuuteen, jolla on yksinkertainen determinismi, parin tai pienen joukon geenipareja., ja jossa todennäköisyyksillä oli suuri merkitys. Genetiikka on vasta-intuitiivinen., Se ennustaa, että ei ole lisääntymiselle emoyhtiön hänen jälkeläisensä, koska jälkimmäinen on genotyypin tulokset lotto erottaminen alleeleja, ennen kuin fuusio kahden vanhemman puolen geenit. Huomattiin, että nämä ovat alleelisia taajuuksia, joissa muunnoksen alleeli esiintyy populaatiossa. Ilmaistuna osuutena tai prosentteina. Populaatiossa olevien geenin alleelien alleelitaajuuksien summa on siis määritelmän mukaan 1. Populaatiogenetiikassa alleelien esiintymistiheydet edustavat populaatio-tai lajitasolla geneettistä monimuotoisuutta., jotka tarttuvat sukupolvien välillä, eivät genotyyppien tai fenotyyppien. Nämä taajuudet ovat enemmän tai vähemmän vakaa sukupolvelta seuraavalle ja tuottaa yli useita takertua filiations yksilöiden välillä, sama genotyyppi frequenciesGenetic rakenne väestöstä. Määritetään alleelisten taajuuksien perusteella (Kuva 4). Populaation genotyyppien jakautuminen, joka liittyy varianssiin, on siten ainoa ennustettavissa oleva tekijä., Noin 1930, kolme teoreetikot Ronald Fisher, JBS Haldane ja Sewall Wright auttoi luomaan käsitystä Mendelin populationPopulation, joiden perinnöllisyys noudattaneet Mendelin lakien mukaisesti. . Tässä yhteydessä kehitys välttämättä associates kolme rakenteellista tasoa: geenin, yksilön ja väestön. Tämä kolme-tie yhdistys voidaan tiivistää seuraavalla kaavalla: väestön kehitys on muutos-alleelia taajuus (gene mittakaavassa, mutta myös ryhmä-asteikko), muutos, joka riippuu lajittelu fenotyyppeihin valinta (yksittäisiä mittakaavassa)., Empiirinen väestön genetiikan tutkimus sitten kehitetty, mutta rajoittui pitkään tietämättömyys, miten kromosomit työ (DNA rakenne oli selvää, vuonna 1953, ja sen sekvensointi alkoi vuonna 1977), ja piti turvautua tutkimuksen harvat näkyvä polymorfismeja, kuten väri leppäkerttu on elytra tai kuori etanat (ks Keskittyä suuri etana keskustelu). Geneetikkojen keskuudessa käytiin paljon keskustelua siitä, olivatko luonnon populaatiot yleensä polymorfisia vai monomorfisia, ja oliko polymorfismi itsessään hyödyllistä vai ei., Tutkimuksen makromolekyyliyhdisteisiin mittakaavassa vaihtelu oli odotettava, kunnes 1966 (proteiineja) ja 1983 (DNA) ennen vastaamista näihin kysymyksiin.
Määrälliset vaihtelut ja periytyvyys
Kun katsomme ympärillemme, näemme fyysiset erot olentoja, jotkut monimutkainen, kuten kasvojen piirteitä, toiset helppo mitata, kuten paino tai korkeus. Maalaisjärki viittaa siihen, että ne ovat osittain tarttuvia, vaikka sitä on vaikea määritellä. Nämä ominaisuudet ovat usein polygenicunder riippuvuus monet geenit. Puhumme polygeenisestä perinnöllisyydestä. Diabetes on polygeeninen sairaus.,, mikä tarkoittaa, että niihin vaikuttavat monet lokukset. Developmental genetics, ja sen soveltaminen vertailu lajien (evo-devo, sillä ”kehitys kehitys”) purkaa web monimutkaisia vuorovaikutuksia, jotka tekevät, että kehityksen aikana kehon soluihin, joissa kaikissa on sama geneettinen matkatavara -, erottaa itsensä epigenesis antaa eri kudoksissa. Vielä on epäselvää, miten monimutkaisia hahmoja rakennetaan. Mitattavissa olevia fenotyyppejä voidaan tutkia biometriikan avulla tilastollisten jakaumien ominaisuuksien perusteella., Yksi arvo hyödyllisiä ominaisuuksia on varianceIn otos yksilöitä, jossa tietty piirre oli mitattu, varianssi on ero välillä tarkoita neliön arvot ja neliön keskiarvo arvoja. Tämä aina positiivinen toimenpide kertoo yksilöiden hajaantumisesta. . Kun useat riippumattomat syyt määrittää vaihtelu linja, niiden varianssit ovat additiivisia, ja niiden summa antaa varianssi line. Jos ne eivät ole riippumattomia, kovarianssien summa lisätään varianssien summaan., Yhteensä varianssi Vt on fenotyyppinen piirre on sellainen, että :
Vt = Vga+Vgd+Vgd+Vgi+Ve
Vga, additiivinen geneettinen varianssi, on summa varianssit, koska jokainen locus muista riippumatta; Vgd, hallitseva geneettinen varianssi, tulokset vuorovaikutus kaksi alleelia samasta locus; Vgi, vuorovaikutus varianssi locuses, tulokset siitä, että locuses saman yksilön vuorovaikutuksessa; Ve, varianssi ympäristö, on oletettu tässä riippumattomiksi geneettinen varianssi.,
geneettinen varianssi valta-asema on yleinen kaksi lasta saman vanhemmat, koska he jakavat valta-aseman vaikutuksia, kun tietyn lokuksen he ovat saaneet samaa alleelia molemmilta vanhemmiltaan, joka tapahtuu heille yksi neljästä locus. He eivät jaa näitä vaikutuksia vanhempiensa kanssa, joten he ovat ”samankaltaisia” toistensa kanssa kuin ne vanhemmat, huolimatta siitä, että he jakavat niin monia geenejä keskenään. Isä tai äiti on varmasti saanut sattumalta samat kaksi alleelia kuin yksi hänen lapsistaan tietyssä lokuksessa., Tämä mahdollisuus riippuu alleelien esiintymistiheydestä populaatiossa. Näemme, että lapsen ja hänen vanhempansa samankaltaisuus tietyn ominaisuuden suhteen on julkista omaisuutta.
yleisemmin kaikki geneettisen varianssin komponentit (Vga+Vgd+Vgi) voivat vaihdella populaatioiden välillä. Jalostukseen linjat, yhteinen kyseessä on sukusiitos, joka pakoputkia geneettinen varianssi: tämän seurauksena piirre voi olla sama keskiarvo kahden populaation, mutta on eri geneettinen vaihtelu. Kasvattajan kyky valita ominaisuus on siis viivalle, ei piirteelle ominainen ominaisuus.,
vanhempien ja lasten suhdetta mitataan heritabiliteetillä h2. Se on määritelty suhde additiivinen geneettinen varianssi yhteensä varianssi :
h2 = Vga/Vt

(2) Korrelaatio 0, jossa koko lasten on riippumaton vanhemmista. Arvokas keskiverto vanhempi on tuottanut välinpitämättömän arvoisen lapsen lasten jakautumisen puitteissa. Pisteiden koordinaatit sijaitsevat ympyrässä; niiden arvon odotus sijaitsee vanhempien akselin suuntaisella viivalla (d)., (3) väli vanhemman ja lapsen välinen korrelaatio välillä 0 ja 1: pisteiden koordinaatit sijaitsevat ellipsillä. Odotus niiden arvo ei ole enää sijaitsevat bisector, koordinaattiakselien (joka on myös pääakselin jakelu ellipsi), mutta regressiolinjan r. Keskimäärin vanhempi arvo on laatinut lapsi-arvo b (välillä 0 ja a) tietty varianssi. On huomattava, että tapauksessa (3) on välimuoto tapaukset (1) ja (2)
Se on osoittanut, eri oletuksia Kuvassa 5., Heritability on kyky määrittää, voidaanko ominaisuus valita. Kuva 5-3 osoittaa, että kasvattaja pystyy kuluessa rajan sukupolvi, liikkua keskimäärin merkin arvo 0 arvo b, jos hän valitsee yksilöiden arvoa. Voimme osoittaa, että b = h2a. Valitse ominaisuus, se on hyödyllistä lisätä periytyvyys suhde standardoimalla ympäristö (vähentää siten ympäristön varianssi), ja asettamalla itsensä ympäristön olosuhteet, jotka suosivat ulkonäkö piirre., Geneetikko Waddington on osoittanut, että luonnossa ympäristön muutos voi siten paljastaa uusia piirteitä, jotka sitten valitaan evoluution aikana.
Viitteet ja huomautukset
kansikuva. Etana kuoret Cepaea nemoralis & Cepaea hortensis. André Künzelmann, UFZ.,
Tämä kaava voi olla kirjoitettu enemmän perinteisellä tavalla, kuten: \pi =\frac{n(n-1))}{2L}\sum _{i=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
koska otos henkilöitä, joille tutkittu piirre oli mitattu, varianssi voidaan määritellä ero tarkoittaa neliön arvot ja neliön keskiarvo arvoja. Tämä aina positiivinen toimenpide kertoo yksilöiden hajaantumisesta.
Tämä on termin tiukassa merkityksessä heritabiliteettiä. Heritabiliteetti laajassa merkityksessä on geneettisten varianssien summan suhde kokonaisvarianssiin., Tämä kaava antaa periytyvyys molempien vanhempien kanssa (”keskimääräinen emo”); arvio, jossa on vain yksi vanhempi antaisi h2/2.
Ympäristön Tietosanakirja Ympäristön Association des Encyclopédies de l ’Environnement et de l’ Énergie (www.a3e.fr), sopimuksen perusteella sidoksissa University of Grenoble Alpes, Grenoble INP, ja sponsoroi ranskan tiedeakatemia.
mainita tämän artikkelin:VEUILLE Michel (2021), Geneettinen polymorfismi ja vaihtelua, Tietosanakirja-Ympäristössä, url : https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
artikkelit Tietosanakirja Ympäristö ovat saatavilla ehtojen mukaisesti Creative Commons BY-NC-SA-lisenssillä, joka sallii jäljentäminen edellyttää: vedoten lähde, ei tehdä kaupalliseen käyttöön niistä, jakaa samat alkuehdot, toistamaan jokaisen uudelleenkäyttö-tai jakelu-maininta tämän Creative Commons BY-NC-SA-lisenssillä.
Vastaa