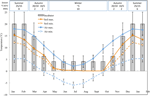
Figure 1. Données mensuelles sur la température maximale et minimale de l’air et du sol dans les Alpes australiennes (moyenne ± STDEV). Les températures de l’Air sont des moyennes de la base de données BOM, les températures du sol ont été mesurées avec des iButtons (voir méthodes). Les barres indiquent les températures expérimentales de l’incubateur (moyenne ± STDEV)., La période équivalente de l’année (saison), le régime de température de l’incubateur (°c, 12/12 h, clair/foncé) et la durée du traitement (semaines) comme indiqué dans le panneau supérieur.
en vue de découvrir des conditions de température proches de celles que connaissent les graines après la dispersion, des données quotidiennes sur la température maximale et minimale du sol ont été recueillies sur le site d’étude (Seaman’s Hut; 36,27°s et 148,17°E, 2030 m A. S. L.) à l’aide d’enregistreurs de données ibutton (n = 10, Embedded Data Systems, USA) placés à 4 cm sous la surface du sol à la base de végétation du 17 janvier au 17 décembre 2012., Nous avons placé des bûcherons à 4 cm pour éviter les perturbations de surface tout en restant représentatifs des conditions auxquelles les graines sont exposées dans le sol; cette profondeur est intermédiaire à ce qui a été utilisé dans les exercices précédents de surveillance de la température du sol alpin (Scherrer et Körner, 2011; Pauli et al., 2015). Comme on le soupçonne, la température moyenne du sol n’est pas descendue sous le point de congélation en hiver (Figure 1). En été, on sait que la température du sol sous végétation suit la température ambiante, alors que la température du sol nu dépassera la température ambiante jusqu’à 15°C (Soil Conservation Service unprecedented records, années 1960-1970).,
collecte et Germination des graines
des graines matures de 54 espèces de 16 familles et 37 genres ont été récoltées entre janvier et avril 2009, 2010 et 2011 (Voir le tableau 1 pour les noms complets et les autorités. Les bons ont été déposés à L’Herbier National Australien, Canberra). Au total, l’espèce représentait plus du quart de la flore des angiospermes australiens présents dans les régions alpines (Costin et al., 2000, Voir tableau 1), bien que beaucoup de ces espèces s’étendent également au-dessous de la limite des arbres., La viabilité de toutes les collections a été estimée avant le semis dans des conditions de germination expérimentales à l’aide de la technique de coloration au chlorure de tétrazolium (TZ) (International Seed Testing Association, 2003). Pour plus de détails sur la collecte et le traitement, voir L’Annexe A dans les documents supplémentaires.
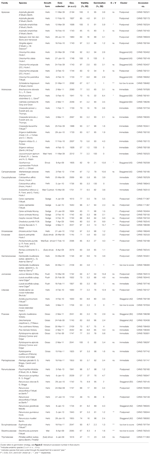
le Tableau 1. Espèce de l’étude, famille et autorité, forme de croissance et détails des collections, pourcentage de viabilité des semences (TZ-estimé, ± S. E.), pourcentage final de germination atteint (± S. E.,), différence statistique entre viabilité et Germination (G < V, testé uniquement dans les cas où la germination était inférieure à la viabilité).
la phénologie de la germination des graines a été étudiée en imitant les régimes de température que les graines alpines connaissent in situ, après la dispersion, dans une progression artificiellement raccourcie des saisons. Les graines ont été imbibées tout au long de l’expérience, ce qui indique un potentiel de germination lorsque l’eau n’est pas limitative., Les données sur la température de l’Air et du sol (Figure 1) ont été utilisées pour guider et valider les régimes de température des incubateurs, bien que les contraintes logistiques du travail avec les incubateurs aient empêché l’incorporation des fluctuations inhérentes aux conditions naturelles. Les tests de Germination ont utilisé huit répliques de 25 graines par collection, semées dans des boîtes de Pétri en plastique de 9 cm de diamètre contenant 1% d’eau-gélose ordinaire. Les boîtes de Pétri ont été scellées à L’aide de Parafilm pour éviter la dessiccation de la gélose, avant d’être placées dans des incubateurs de germination (Thermoline Scientific, Melbourne, NSW, Australie)., La moitié des plats étaient enveloppés dans du papier d’aluminium pour exclure la lumière et fournir une indication des besoins en lumière pour la germination. Chacune des quatre répliques par traitement à la lumière et par collection a été placée sur une étagère différente dans le même incubateur et l’emplacement dans cette étagère a été réorganisé chaque semaine. Toutes les répliques de chaque collection ont été déplacées à travers les thermopériodes simulées suivantes: 8 semaines à 25/15°C (Température jour/nuit) → 2 semaines à 20/10°C → 2 semaines à 10/5°C → 10 semaines à 4-5°c constant → 2 semaines à 10/5°C → 2 semaines à 20/10°C → 6 semaines à 25/15°C., Ainsi, un cycle de toute l’expérience a imité des températures rappelant l’été, le début de l’automne, la fin de l’automne, l’hiver, le début du printemps, la fin du printemps et l’été, et a duré 32 semaines au total. L’année abrégée a été jugée raisonnable parce que les accessions avaient généralement cessé de germination dans un traitement donné avant d’être déplacées à la « saison”suivante., Une photopériode lumière/obscurité de 12/12 h a été fournie tout au long par des tubes fluorescents (> 90 µmol m−2 s−1 au centre de chaque étagère) et, au printemps, en été et en automne, la lumière a coïncidé avec la période de température chaude de la journée. La Germination, définie comme l’émergence de radicules de plus de 1 mm, a été notée tous les 7 jours, et les graines germées et les plantules ont ensuite été retirées des plats. La Germination des plaques enveloppées de papier d’aluminium a été évaluée comme ci-dessus, mais sous une très faible lumière verte.,
Après un cycle expérimental (32 semaines), la plupart des collections ont été interrompues (voir les informations supplémentaires pour les exceptions), après avoir montré une germination nulle pendant au moins 2 semaines à ce moment. Après la fin, toutes les graines intactes restantes ont été disséquées au scalpel au microscope. Les graines avec un endosperme ferme et frais et un embryon ont été jugées viables et les graines vides d’un embryon ont été déduites du total lors du calcul du pourcentage de germination pour chaque collection., Nous avons calculé un indice des besoins en lumière pour la germination pour chaque espèce en tant que fraction de la germination dans le traitement à l’obscurité au moment où les percentiles cumulatifs de germination de 50 et 95% ont été atteints dans le traitement alternant lumière/obscurité (voir ci-dessous).
corrélats écologiques de la stratégie de Germination
Nous avons compilé une base de données des traits fonctionnels des espèces et des mesures géographiques à partir des données publiées pour évaluer les corrélations avec la stratégie de germination., La base de données comprenait les caractères continus suivants: masse moyenne des graines, besoin de lumière pour la germination (voir ci-dessus), surface foliaire spécifique (surface fraîche/masse sèche), surface foliaire individuelle, poids sec individuel des feuilles et hauteur à maturité. Les élévations minimales, moyennes et maximales de la collection pour l’espèce ont été déterminées à partir de tous les enregistrements d’herbier dans L’Herbier virtuel australien, et l’altitude à laquelle les collections de semences spécifiques utilisées dans l’étude ont été effectuées a également été incluse comme caractères continus. La forme de croissance a été notée comme l’une des trois catégories: graminoïde, herbe ou arbuste., Nous avons également inclus un trait binaire: si les graines étaient endospermiques ou non.
Les données sur les caractères ont été recueillies auprès de diverses sources, notamment L’Herbier virtuel Australien (http://avh.chah.org.au/), le Registre National D’information sur les spécimens D’Herbier Australien (https://www.anbg.gov.au/cgi-bin/anhsir), la flore alpine de Kosciuszko (Costin et al., 2000), La Flore De La Nouvelle-Galles du Sud (Harden, 1990) et la morphologie interne comparative des graines (Martin, 1946). La masse des graines et la surface foliaire spécifique ont été obtenues à partir de collections individuelles au champ utilisées dans les essais de germination., La masse des graines a été obtenue en pesant quatre lots de 10 graines. L’als a été déterminée sur la base de cinq feuilles entièrement élargies provenant d’individus distincts par espèce. Ceux-ci ont été recueillis sur le terrain, scannés sur un scanner à plat, séchés à 60°C à un poids constant et pondérés à 0,000 g près. lorsque les graines étaient insuffisantes ou que la surface foliaire spécifique n’était pas disponible, les données ont été obtenues à partir d’autres collections ANBG de la même espèce ou d’autres sources de données (s. Venn, données non publiées)., La question de savoir si une graine était endospermique ou non a été déterminée à partir de dessins de la graine dans les dossiers de la Banque de semences ANGB en consultation avec le personnel de la Banque de semences ANBG. La base de données complète et les sources sont disponibles sur demande auprès des auteurs.
analyse des données
Les Patrons de Germination ont été évalués visuellement et à l’aide d’une analyse en grappe dans PC-ORD Pour Windows, version 6 (MJM Software Design, Gleneden Beach, OR, USA). L’analyse a utilisé une matrice de distance euclidienne et un algorithme de jointure de voisin le plus proche., Les points de données pour l’analyse en grappes étaient le temps (semaines) jusqu’à des percentiles de germination cumulatifs spécifiques (T20, t40, T50, T60, t80 et T95%) pour chaque collection (moyenne pour les 4 plats). Ainsi, une collection qui a germé tôt dans l’expérience aurait des temps courts aux percentiles jusqu’à t80% par exemple, alors qu’une collection qui a reporté la germination jusqu’à ce que les températures augmentent après l’hiver simulé aurait des temps longs à ces percentiles., Des schémas de germination plus complexes, tels qu’une germination lente et régulière ou des rafales de germination, pourraient également être détectés et seraient indiqués par des temps régulièrement espacés ou des temps regroupés au début et à la fin de l’expérience, respectivement. Cette méthode a été utilisée pour imposer une évaluation objective du schéma de germination, indépendamment de la température en soi.,
Une analyse unidirectionnelle de la variance (ANOVA) a été effectuée pour les plaques répliquées de chaque collection afin d’évaluer s’il y avait une différence significative entre le pourcentage de viabilité (tel que déterminé par le test TZ) et le pourcentage final de germination. D’autres modèles linéaires généraux ont été utilisés pour évaluer si les grappes résultant de l’analyse des grappes différaient en termes de viabilité ou de pourcentage de germination final.
Les Associations entre les caractères écologiques et les clusters de la stratégie de germination ont été testées de deux façons., Premièrement, nous avons testé les différences moyennes entre les grappes en utilisant ANOVA avec cluster comme effet fixe et les corrélats écologiques comme variables de réponse, ou en utilisant des tests g répliqués pour l’endospermie et la forme de croissance. Deuxièmement, parce que la germination et d’autres traits fonctionnels sont très susceptibles d’être influencés par l’histoire évolutive, nous avons évalué ces corrélations en utilisant des régressions phylogénétiques. Une phylogénie a été construite pour l’espèce à l’aide de Phylocom Phylomatic et de l’emballage de bladj dans R (Webb et al., 2008; R Équipe De Base, 2013)., Nous avons évalué la présence de la structure phylogénétique dans les variables en utilisant le phylosig ou phylo.d (variables discrètes ou binaires) Fonction Du paquet Phytools dans R (Revell, 2012). Des régressions phylogénétiques ont été réalisées à l’aide de la fonction pgls chez Caper (Orme et al., 2013). Les Analyses ont été effectuées de manière structurée, reflétant le regroupement révélé dans PC-Ord (voir ci-dessous)., Initialement, nous avons inclus quatre stratégies de germination (catégories différées, immédiates et décalées dans chacune des catégories précédentes), puis trois (regroupant toutes les espèces décalées et comparant à différées et intermédiaires) et enfin nous avons évalué les différences entre les deux stratégies les plus larges, différées et intermédiaires.
résultats
viabilité des graines par rapport à la Germination finale
dans l’ensemble, la viabilité des graines et le pourcentage de germination finale étaient élevés. La viabilité moyenne estimée par TZ dans toutes les collections était de 80 ± 1.,7%, plus des deux tiers des collections présentant une viabilité supérieure à 75% (Tableau 1). Il n’y a pas eu de différence dans la qualité des collectes au cours des 3 années (la viabilité Moyenne en 2009, 2010 et 2011 était de 79 ± 2,8, 80 ± 2,7 et 81 ± 3,7%, respectivement), ce qui indique que la Banque de semences récoltées en 2009 n’a pas diminué leur viabilité.
une germination qui dépassait ou n’était pas significativement inférieure à la viabilité estimée par TZ a été atteinte pour 45 des 54 espèces (ANOVA unidirectionnelle: P> 0,05, Tableau 1)., Sur les neuf autres espèces, la germination finale de quatre espèces était très faible (< 5%), malgré une bonne viabilité (env. 55-94%), et nous n’avons donc pu tirer aucune conclusion sur les stratégies de germination de ces espèces. L’échec de la germination pourrait refléter soit que nos traitements n’ont pas atténué la dormance, soit que les graines n’étaient pas complètement matures au moment de la collecte. Les tests de coupe à la fin de l’expérience ont indiqué que, dans tous les cas, la majorité des graines non germées contenaient un embryon sain et imbibé., Fait intéressant, 34 espèces ont obtenu une germination supérieure à la viabilité estimée par TZ (Tableau 1), ce qui suggère que le test TZ sous-estimait généralement la viabilité des graines.
grappes de Germination
l’analyse des grappes a révélé deux grappes principales (A et B), avec une sous-structure dans chacune (Figure 2). Pour la majorité des espèces (n = 21 espèces de 10 familles), la germination a été reportée à une période de températures hivernales (5°C jour/nuit) ou jusqu’à ce que les températures soient élevées à 10/5°C après l’hiver (A1 et A1:hiver, Figures 2, 3A)., Alternativement, la germination a commencé immédiatement à 25/15°C (n = 17 espèces, six familles, vues dans le groupe B1, Figures 2, 3b). Pour la majorité de ces espèces, le taux de germination était rapide; 50% de la germination totale (t50) s’est produite en moins de 3 semaines (figure supplémentaire 1).
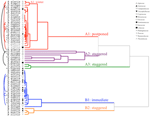
la Figure 2. Analyse en grappes dendrogramme basé sur le temps à percentiles de germination cumulés(20, 40, 50, 60, 80, et 95%). Espèce indiquée par l’initiale du genre et la première partie du nom de l’espèce. Les astérisques marquent les collections qui ont été mises en banque avant les tests., Des arcs fins relient des collections en double pour comparer l’effet de la banque sur la germination. Les symboles désignent différentes familles (voir légende). Les Collections présentées en texte gris sont celles qui ont été exposées à deux cycles de l’expérience.
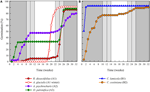
la Figure 3. Pourcentage cumulatif de germination d’une espèce représentative de chaque grappe de germination. (A) grappe A, espèce qui a reporté la germination, (B) grappe B, espèce qui a germé immédiatement., Notez que les schémas de germination échelonnés sont imbriqués dans les grappes ci-dessus. Les régimes de température de l’incubateur sont représentés par l’ombrage (voir la Figure 1).
dans chacune de ces grandes classes, cependant, il y avait des sous-grappes claires reflétant une germination échelonnée dans le temps, c’est-à-dire qu’une partie du lot de graines a germé avant l’exposition aux températures hivernales, tandis que le reste a germé après l’hiver. La germination échelonnée a eu lieu chez 13 espèces de six familles et s’est produite principalement lorsque les températures diurnes dépassaient 10°C (Figures 3A,B)., Ces germinateurs décalés ont été divisés en trois groupes dans l’analyse en grappes. Le groupe A2 avec sept collections a montré une germination importante avant et après la période de température froide. Lorsque la germination a cessé avant que les températures ne changent, nous avons conclu que l’absence de germination supplémentaire avant l’hiver n’était pas due à la température (A3). En revanche, la germination d’une grande partie du groupe A2 ne semblait être arrêtée que lorsque les températures étaient réduites à 10/5°c., Le groupe B2 contenait quatre autres espèces qui ont montré une germination importante au début de l’expérience et qui ont ensuite atteint une proportion relativement faible de leur germination totale après le retour des températures au-dessus de 5°C (Figure 3B).
par rapport à la variation relativement mineure entre les espèces au sein des grappes A1 et B1, les grappes décalées montrent également une différenciation beaucoup plus grande entre les espèces. Les courbes individuelles pour toutes les espèces sont présentées dans les Figures supplémentaires 1i-vii.pour plus de détails sur la stratégie de germination, voir L’Annexe B dans les documents supplémentaires.,
corrélats écologiques de la stratégie de Germination
de tous les corrélats écologiques que nous avons considérés, seuls ceux directement associés aux caractères des graines étaient significativement corrélés à la stratégie de germination, que nous ayons évalué la corrélation au niveau de deux grands groupes (germination différée par rapport à la germination immédiate), ou au niveau de trois ou quatre groupes, ou que nous ayons tenu compte de la structure phylogénétique ou non (Tableau 2)., Les espèces dont la germination a été reportée avaient des graines plus lourdes et étaient plus susceptibles d’être endospermiques que celles des espèces qui ont germé immédiatement (Figure 4). Les espèces à germination échelonnée étaient intermédiaires en masse de graines et en endospermie. Cependant, toutes les caractéristiques des graines n’étaient pas corrélées avec la stratégie de germination: il n’y avait pas de corrélation entre les besoins en lumière pour la germination des graines et la stratégie. Contrairement à nos attentes, les espèces ayant une surface foliaire spécifique plus élevée (ce qui pourrait indiquer des taux de croissance plus élevés) n’étaient pas plus susceptibles de présenter une germination immédiate., De même, les espèces ayant une altitude moyenne plus élevée ou des plages d’altitude plus petites n’étaient pas plus susceptibles de présenter une germination différée.

le Tableau 2. L’étude de la structure phylogénétique et les corrélats de la stratégie de germination.
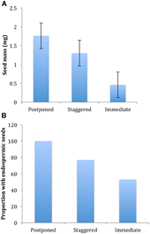
la Figure 4., Différences dans (A) la masse moyenne des graines et (B) la proportion d’espèces avec des graines endospermiques entre les espèces avec des stratégies différées (grappe A1), décalées (combinaison des grappes A2, A3 et B2) et de germination immédiate (grappe B1). Les différences entre les stratégies sont significatives à P ≤ 0,05, voir le tableau 2.
Discussion
la Germination dans les habitats alpins était historiquement considérée comme contrôlée sur le plan environnemental, la neige hivernale isolant les graines contre les indices de germination potentiels et rendant la dormance inutile (Billings et Mooney, 1968)., Cependant, nos résultats soutiennent des preuves plus récentes de mécanismes de dormance communs chez les espèces alpines en Australie (Hoyle et al., 2013; Sommerville et coll., 2013) et ailleurs dans le monde (Cavieres et Arroyo, 2000; Schutz, 2002; Shimono et Kudo, 2005; Mondoni et coll., 2009; Schwienbacher et coll., 2011)., La dormance semble jouer un rôle important dans le contrôle du moment in situ de la germination de nombreuses espèces alpines australiennes, agissant pour retarder, reporter ou ralentir le taux de germination et donc potentiellement conférer des stratégies de régénération sans risque dans un environnement difficile et variable. Le plus frappant de nos résultats de germination était peut-être la proportion d’espèces pour lesquelles la stratégie de germination variait au sein d’une collection de graines pour une espèce donnée, ce qui suggère que les caractéristiques dormantes et non dormantes étaient présentées au sein de la même collection de graines de ces espèces., Notamment, les stratégies de germination étaient généralement très conservées au sein des familles et étaient également corrélées avec des éléments de l’anatomie des graines: masse et endospermie. Les stratégies de Germination étaient cependant indépendantes des autres caractéristiques fonctionnelles écologiquement significatives de la plante mature.
variation de la stratégie de Germination
Les Graines de près de la moitié des espèces étudiées semblaient incapables de germer avant d’être exposées à une période fraîche et humide (constante 5°C), ce qui suggère que la stratification froide atténuait un mécanisme de dormance physiologique., Le report de la germination au printemps suivant peut permettre aux semis d’éviter de s’établir au-dessus ou avant l’hiver rigoureux, tout en optimisant la courte saison de croissance à venir. En revanche, la germination opportuniste des espèces qui ont germé immédiatement peut fournir un avantage sélectif lorsque le risque de mortalité des semis en hiver est faible, en permettant aux plantes de fleurir plus tôt le printemps suivant ou à une taille plus grande (Donohue, 2002).,
à ce jour, il y a eu peu de preuves publiées de variabilité intra-spécifique dans les mécanismes de dormance au sein des collections de semences alpines, ce qui expliquerait les stratégies de germination échelonnées observées ici. La germination échelonnée dans le temps peut également s’expliquer par des niveaux variables de maturité des graines chez les individus de la population au moment de la collecte, et/ou pourrait refléter une stratégie dimorphique au sein d’une seule plante associée à la position sur la plante ou au moment du développement.,
la couverture de pari évolutive est souvent évoquée pour expliquer la stratégie diversifiée dans les caractéristiques de germination des graines (Simons, 2011), comme celle démontrée par les collections avec germination échelonnée. Toutefois, si une stratégie diversifiée se traduit à la fois par une amélioration de la condition physique moyenne et une réduction de la variance (par exemple, en réduisant la concurrence entre frères et sœurs), elle ne constitue pas une couverture de pari. De même, si la diversification apparente reflète les différences entre les individus, et non au sein de ceux-ci, elle ne constitue pas une couverture de pari (Starrfelt et Kokko, 2012)., Les études sur les schémas de dispersion des semis alpins et la tolérance au gel et à la fonte des neiges, jumelées à des études sur les déterminants de la variation de la stratégie de germination au sein des espèces, peuvent aider à expliquer la phénologie de germination des espèces présentant une stratégie échelonnée et peuvent indiquer si elles sont susceptibles de représenter des stratégies de couverture de pari.,
corrélats écologiques de la stratégie de Germination
notre examen de la relation entre la stratégie de germination et d’autres caractères écologiquement importants a démontré que les caractères de germination étaient corrélés avec d’autres caractères de la graine, mais pas avec des caractères végétatifs. Sommerville et coll. (2013) ont constaté que les herbes alpines australiennes avec des graines non endospermiques étaient plus susceptibles d’être non dormantes à la dispersion. Nos travaux confirment cette constatation et indiquent en outre que les graines plus petites sont plus susceptibles d’être parmi les germinateurs immédiats. Bien que Sommerville et coll., (2013) n’ont pas avancé pourquoi l’endospermie peut être associée à la dormance, nous suggérons que le placement de réserves dans les cotylédons pourrait améliorer le taux de croissance précoce et l’établissement Chez les espèces en germination immédiate. Nos résultats n’indiquent aucune corrélation entre les besoins en lumière et la dormance, bien que les besoins en lumière varient d’une espèce à l’autre. Il existe une variété d’autres caractéristiques de la graine qui atténuent le risque d’extinction dans des environnements variables, par exemple les mécanismes de dispersion des graines et la longévité des graines dans la Banque de graines du sol (Voir Tielborger et al., 2012 pour un examen)., À ce stade, nous ne pouvons pas dire si ces caractères montreraient des corrélations avec la stratégie de germination ou non.
nous avons également demandé si la stratégie de germination serait corrélée avec des caractères non reproducteurs d’importance écologique, tels que la surface foliaire spécifique ou la hauteur de la plante (Westoby, 1998; Wright et al., 2004). En particulier, nous avons postulé que les espèces à germination immédiate pourraient montrer des valeurs de caractères indiquant des taux de croissance plus rapides (en utilisant L’als comme indicateur) qui pourraient être nécessaires pour l’établissement dans la courte période entre la dispersion et le début de l’hiver., Cependant, nous n’avons trouvé aucune corrélation de ce type. La question de savoir si les traits physiologiques et de croissance des plantules aux premiers stades de l’établissement pourraient montrer des modèles différents de ceux des traits végétatifs adultes est une question qui mérite d’être examinée, mais qui dépasse la portée du présent travail.
enfin, nous avons examiné si les espèces provenant d’altitudes plus élevées où les saisons de croissance sont généralement plus courtes pourraient être plus susceptibles de présenter une dormance, mais il n’y avait aucune association entre la plage d’altitude, les élévations de collecte ou l’altitude moyenne de la collecte pour l’espèce dans son ensemble et la stratégie de germination., Ensemble, ces résultats indiquent que, bien que l’évolution de la stratégie de germination soit étroitement liée aux caractéristiques écologiques importantes de la graine, elle comprend un axe largement indépendant des caractéristiques végétatives ou distributionnelles.
Implications pour les communautés de plantes alpines dans un climat changeant
Les écosystèmes alpins australiens, comme les zones alpines du monde entier, sont menacés par le changement climatique, combiné à des changements dans la fréquence des incendies, les schémas d’utilisation des terres, l’afflux d’espèces envahissantes et les impacts de la fréquentation humaine accrue., Le réchauffement associé au changement climatique se produit plus rapidement au-dessus de la limite des arbres qu’à des altitudes plus basses, et les régions alpines devraient continuer à connaître un réchauffement supérieur à la moyenne à l’avenir (Kullman, 2004). La capacité de régénération continue des semences dans de nouvelles conditions est susceptible de jouer un rôle important dans la réponse des plantes alpines au climat et aux changements qui y sont associés. En outre, les régimes de perturbation modifiés nécessiteront probablement un rôle accru de restauration et de remise en état dans la région., Les informations fournies ici sont donc cruciales pour la conservation et la gestion des systèmes alpins en mutation; elles peuvent aider à éclairer nos prédictions sur la façon dont les espèces étudiées peuvent être affectées, constituer la base des plans de propagation des semences de ces espèces et être utilisées pour guider les futures recherches sur les stratégies de germination des semences alpines,
Les changements de température sont susceptibles d’affecter le statut de dormance des graines avant et après la dispersion, modifiant et/ou perturbant les stratégies de germination et entraînant des changements dans la phénologie actuelle de la germination (Hoyle et al., 2008b). Pour les plantes alpines, le réchauffement climatique peut conduire à un déplacement de l’émergence du printemps à l’automne, entraîné principalement par des changements dans le statut de dormance des graines et entraînant des implications majeures pour les espèces actuellement adaptées à l’émergence au printemps (Mondoni et al., 2012)., Des recherches supplémentaires sur la façon dont les changements de température que subissent les graines après la dispersion, la durée réduite de la couverture neigeuse et la fréquence accrue des événements de température extrêmes affecteront la germination sont nécessaires afin de prédire comment les espèces en dormance réagiront au changement climatique. Pour les espèces alpines qui germent en automne, avant le gel et les chutes de neige, nos résultats soulèvent la question de savoir comment les semis de ces espèces vont faire face à la réduction prévue des précipitations et à l’augmentation de la fréquence des épisodes de gel saisonnier A.,
enfin, nos résultats soulèvent des questions importantes concernant la variation au sein des espèces, en particulier en association avec la prédiction de l’impact du changement climatique ou la sélection des graines à utiliser pour la restauration. Les stratégies de Germination peuvent varier au sein de l’espèce en fonction de la population, du temps de floraison, de la masse des graines, de l’investissement parental, du climat, de l’altitude et/ou de l’environnement maternel, et cette variation peut refléter des facteurs génétiques, épigénétiques et/ou environnementaux (Cochrane et al., 2015)., L’augmentation de l’altitude explique la variation de la germination et / ou du statut de dormance de plusieurs espèces alpines (Billings et Mooney, 1968; Dorne, 1981; Cavieres et Arroyo, 2000), mais voir (Hoyle et al., 2014). L’état de dormance réduit peut résulter de la fin précoce du développement des semences résultant de stress environnementaux (Steadman et al., 2006; Hoyle et coll., 2008b)., L’examen de la variation des stratégies de germination au sein des espèces peut révéler la possibilité pour les espèces de faire face aux changements climatiques et de s’y adapter, jouant ainsi un rôle important dans la détermination de la survie future et de la répartition des espèces et fournissant des informations importantes pour la gestion et la conservation.
Conclusions
notre approche a révélé une présence généralisée de dormance dans la flore alpine australienne et démontre que les stratégies de germination varient au sein d’une collection de graines, ainsi qu’entre les collections., De plus, nous avons montré que bien que la stratégie de dormance soit conservée phylogénétiquement et corrélée à la masse des graines et à l’endospermie, elle est largement indépendante des traits végétatifs et des caractéristiques de l’aire de répartition. Bien qu’on ne puisse pas extrapoler directement à partir de nos résultats comment le climat et les changements associés affecteront la flore alpine, ces résultats peuvent servir de base à la conception de plans de propagation et à d’autres expériences sur l’impact des changements de régimes climatiques., En particulier, nous préconisons des recherches plus poussées pour comprendre les causes de la variation intra-espèce de la stratégie de germination et le rôle que cela peut jouer dans la capacité des espèces à résister aux changements climatiques prévus.
accessibilité des données
Les données complètes sur la germination et la base de données des caractères seront déposées sur DRYAD et auprès de L’Atlas of Living Australia.
déclaration de conflit d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de toute relation commerciale ou financière pouvant être interprétée comme un conflit d’intérêts potentiel.,
Remerciements
Nous reconnaissons le financement de L’Australian Research Council (LP0991593) et du Australian National Botanic Garden et de ses amis. Nous remercions J. McAuliffe, S. Fethers et S. Lawatsch de L’ANBG. Nous remercions également M. Holloway-Phillips, M. Cardillo et S. Cunningham pour leurs contributions sur les versions antérieures du manuscrit et pour leurs conseils analytiques et S. Venn pour l’accès aux données sur les traits.
documents supplémentaires
Foley, M. E. (1992). Effet des sucres solubles et gibbérellique-acide à briser la dormance des extraits de la folle Avoine (Avena fatua) embryons., Les Mauvaises Herbes De La Sci. 40, 208–214.
Google Scholar
Durcir, G. J. (1990). La flore de Nouvelle-Galles du Sud. Sydney, NSW: Jardins Botaniques Royaux.
Google Scholar
l’International Seed Testing Association. (2003). Feuilles de travail sur les tests de tétrazolium. Bassersdorf: ISTA.
Kullman, L. (2004). Le visage changeant du monde de ski alpin. Glob. Changer Newsl. 57, 12–14.
Google Scholar
Laisser un commentaire