
a genetikai polimorfizmus a DNS alternatív állapotainak létezése, meghatározva a szervezet magasabb integrációs szintjeinek variációját. Különböző genommódosítások (mutációk) vannak. A leginkább vizsgált nukleotid szubsztitúciók a kódolási és szabályozási régiókban.
definíció
a genetikai polimorfizmus egy populációban több alternatív DNS-állapot vagy allél létezése, a genomban meghatározott helyzetben, vagy a gén lokuszpozíciója a kromoszómán., A populációs genetikában homológ gének halmaza (homológiai osztály). Két gén homológ, ha megfelelnek a meiózisnak. Többes szám: loci.. Ennek a meghatározásnak számos aspektusa van:
(1) Először is, a tulajdonságot a kromoszómáknak kell hordozniuk, és átvihetőnek kell lenniük.
(2) ezután az alléloknak homológnak kell lenniükkét kromoszómát vagy két gént homológnak kell tekinteni, ha egyeznek, és kölcsönösen kizárják egymást a meiózisból. a genomban betöltött helyzetükre, amely kizárja bizonyos típusú variációkat, amelyeket később tárgyalunk.,
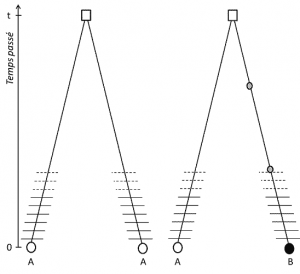
(3) De mivel a tulajdonság átvihető, a pozíció homológia azt is jelenti, hogy az allélok származás szerint homológ; ha különböznek egymástól, akkor ezért mutáció (legalább) történt az egyik sorban, amely összeköti őket az utolsó közös ősükkel (1.ábra).
(4) végül a genetikai polimorfizmus meghatározható a DNS-t alkotó legkisebb egység skáláján: a nukleotid helyén., Ennek eredményeként minden nukleotid variáns meghatározhatja a biológiai szerkezet magasabb szintjeinek – az egyén génjének, fehérjéjének és fenotípusának-polimorfizmusát, amely olyan skálává válik, amelyen a genetikai polimorfizmus leírható (2.ábra).
Vegyük az ABO vérrendszer példáját,amely megfelel az antitest vagy limfocita receptor által elismert antigén motivációinak. Epitóp vagy antigén determináns néven is ismert. Ugyanazon antigénnek több epitópja is lehet (azonos vagy eltérő), így változatos immunválaszt válthat ki., részt vesz a vérátömlesztés során fellépő kompatibilitásban. Ez az emberi populációkban jelen lévő genetikai polimorfizmus esete, amely magában foglalja az ABO-glikozil-transzferázenzim fehérjét, amely lehetővé teszi a cukrokat tartalmazó maradékok fehérjékre történő átvitelét. Az ABO rendszer esetében a glikozil-transzferáz A és B indukálja az egyént az A vagy B csoportba. ha mindkét glikoziltranszferáz jelen van, az egyén az AB csoportba tartozik. amelynek három allélja van, A, B és O. az egyénnek lehetnek (AA), (AO); (BB), (BO); (OO) vagy (AB) genotípusai., Fenotípusa ezután,,;, vagy, ahol azt látjuk, hogy az A és B “uralja” O-t (azaz kifejezése maszkolja az O-t), és hogy egymás között dominálnak (az A és B közötti heterozigóta felismerhető kifejezéssel rendelkezik ).
a polimorfizmus először a fehérjét kódoló Lokus DNS-szekvenciájának skáláján írható le. Egyes polimorfizmusok “szinonimák”, azaz nem változtatják meg a fehérje aminosav-szekvenciáját; gyakran a legtöbb polimorfizmus. A többiek módosítják az aminosavakat, és” helyettesítő ” polimorfizmusoknak nevezik őket., Az ABO-glikozil-transzferáz helyettesítő polimorfizmusai két típusból állnak: az aminosavat megváltoztató polimorfizmusok, de nem az antigén egység (nem zavarják az egyének ABO-fenotípusát); és más helyettesítő polimorfizmusok, amelyek meghatározzák az egyén megfigyelhető jellemzőinek ABO-fenotípusát..
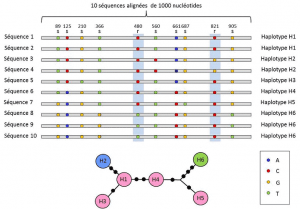
a nukleotid és az allél szintek között a genetikai elemzések figyelembe veszik a leírás közbenső szintjét: a változó helyek lineáris elrendezését a lokuson. Ezek az elrendezések, az úgynevezett haplotípusokkülönböző Loki allélcsoportjai, amelyek ugyanazon a kromoszómán helyezkednek el, és általában együtt továbbítják őket. A haplotípus az angol haploid genotípus vagy haploid genotípus összehúzódása., Minden olyan gén, amely ugyanazon a kromoszómán található, és amelynek alléljai a meiózis során elkülönülnek egymástól, haplotípust képeznek. Ezeket a géneket “genetikailag összekapcsoltnak” nevezik, hasznosak az evolúciós genetikában, mert lehetővé teszik az allélok közötti genealógiai kapcsolatok megtalálását (2.ábra).
nukleotid polimorfizmusok is léteznek intergén régiókban (a kódolási régiókon kívül). Egyesek befolyásolják azokat a területeket, ahol a génexpresszió szabályozott, ezért fenotípusos expresszióval rendelkeznek. Másoknak nincs ismert hatásuk, és “csendesnek”nevezik őket.,
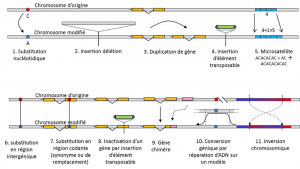
a nukleotid szubsztitúciók mellett egy adott típusú mutáció polimorfizmusa, a mikroszatelliták egy gyakran nem kódoló DNS-fragmens hosszának változásaira utalnak (3.ábra) a nukleotidok rövid ismételt szekvenciájában, például tgtgtgtgtgtg ismétlések számának változása miatt.
nem minden genetikai variáció tartozik a genetikai polimorfizmus definíciójába, mivel az alternatívák homológiáját igényli (3.ábra)., Így vannak olyan ismétlődő szekvenciák, amelyeket egy gén duplikálásával nyernek, amelyekhez két ismétlés pozíciónként nem homológ. Gyakran nem beszélhetünk a transzpozálható elemek homológiájáról sem, mert általában megváltoztatják a pozíciójukat a sejtgenerációk során, szaporodhatnak és behatolhatnak a genomba. Két transzpozálható elem homológiája ezután nem határozható meg. Másrészt ugyanazon hely két kódolási szekvenciája, amelyek közül az egyik sértetlen, a másik pedig egy átültethető elem behelyezésével inaktivált, jól megfelel a homológiai helyzetnek., Egy nap, akkor valószínűleg úgy beszél, arról, hogy polimorfizmus a módosítások úgynevezett “epigenetikus” kromoszóma, amelyek néha továbbított több cella generáció, somaticQualifies nem-reproduktív sejtek, vagy soma. A szomatikus mutáció, amely egy szomatikus sejt génjét érinti, eltűnik a hordozó egyénével. vagy germinálisminősíti az ivarsejteket. A csíravonal mutáció továbbítható utódainak., mint például a metiláció. A populációk fejlődésében betöltött jelentőségüket továbbra is értékelni kell., A genetikai polimorfizmus fogalmának korlátozása bizonyos variációs kategóriákra-lényegében nukleotid szubsztitúciókra-az utóbbi nagy hasznosságából származik a természetes populációk történetének feltárásában. Az evolúció matematikai modellezésére alkalmasak.
a polimorfizmus szó “több formát” jelent. Ellenzi a monomorfizmust, ami a variáció hiányát jelzi. A biológia szókincsében ezt az ellenzéket már régóta használják a genetika előtt, hogy jelezzék többféle egyén együttélését ugyanabban a fajban, például, kaszt polimorfizmus (királynő és munkások) a társadalmi rovarokban; szezonális polimorfizmus (szőrzetváltozások) egyes sarkvidéki emlősökben. Ezek az esetek nem tartoznak a genetikai polimorfizmus, egy újabb fogalom, szűkebb és pontosabb jelentéssel. A folyamatos variációk (pl. méretváltozások) szintén nem tartoznak a polimorfizmus kategóriájába, mivel nem mutatnak külön alternatívákat. A méretet befolyásoló locus azonban ebbe a definícióba tartozik., Az alábbiakban tárgyaljuk a leginkább vizsgált genetikai variációkat, mivel ezek a legfontosabbak az evolúció, a nukleotid szubsztitúciók szempontjából, majd megvitatjuk a fenotípusok variációját.
mérés
nem elég azt mondani, hogy egy Lokus többé-kevésbé változó anélkül, hogy kvantitatív becslést adna erre az ítéletre. A kutató különböző skálákon tanulmányozhatja a variációt. Ha csak egy fehérje allélja érdekli, akkor megméri az allél sokszínűséget, amelyet a ” H ” jelképez. Ha érdekli a DNS-sokféleség, meg fogja mérni a nukleotid sokféleségét, amelyet a ” π ” szimbolizál.,
az allél sokszínűség H úgy definiáljuk, mint a valószínűsége rajz két különböző allélok során két mintavétel replacementPerforming egymást követő döntetlen a szállítási p tokenek egy urn tartalmazó n tokenek, azt jelenti, hogy egy első token, olvasás értékét, amivel vissza az urn, hogy egy második token, olvasás értékét, amivel vissza az urn, stb, amíg a pth token. Ez azt jelenti, hogy P objektumokat választanak az n között ismétléssel (ugyanazt az objektumot többször is kiválaszthatja), valamint sorrendben (az objektumok kiválasztásának sorrendje fontos)., Az egymást követő húzások száma tokenekkel az n között: n × n × n × … × n = np.. Ha a PI-t az I. rang alléljának gyakoriságának nevezzük, megmutatjuk, hogy a kétszer azonos allél kiválasztásának valószínűsége F = Σ pi2. A minta allél sokfélesége ezután kiegészíti az 1-et, azaz:
H = 1 – Σ pi2 (1)
Ez mind a fehérje allélokra, mind a haplotípusokra vonatkozik. Ezt haplotipikus sokféleségnek nevezik. Heterozigóta is nevezhető, mert diploid locus esetén a heterozigóták várható gyakoriságát adja.,
a nukleotid sokfélesége π az egyes nukleotidok skáláján h-val egyenértékű. Ezt a nukleotid-különbségek számának átlagaként számítják ki egy kettővel vett mintában (δij), a DNS-fragmentum hosszával osztva a nukleotidok számára (L).
π = átlag (δij)/L (2) (lásd referencia )
Ez az érték fajonként eltér. Az emberi genom kódoló részeiben a populációból véletlenszerűen kiválasztott két kromoszóma átlagosan ezer nukleotiddal különbözik., A gyümölcslégyben (Drosophila melanogaster)ez a különbség körülbelül száz. A légy tehát tízszer több változó, mint az ember. Amikor ezek az értékek kapcsolódnak a méret a kódoló régiók (körülbelül 15,500 gének a Drosophila, legalább 22,000 az emberek), vagy akár a genom szekvenciát jelent (140 millió bázispár per haploid genom legyek, körülbelül 3,200 millió vagy hússzor több emberben) száma, a polimorf oldalak csillagászati, majd eredménye, hogy senki sem született a szexuális töltetű generációs bármely faj, fog genetikailag azonos egy másik múltban, a jelenben vagy a jövőben., A DNS polimorfizmus információs ereje óriási. A helyszínelőkben a nyomozók tizenhat mikroszatellita lokusból azonosítanak minden gyanúsítottat.
történelem
a variáció kifejezés Darwin “A fajok eredete” című könyvének első két fejezetének címében található (1859). Darwin akkor vezette be ezt a fogalmat a természettudományokba, amikor a biológiai öröklődés törvényei rejtély maradtak, és egy másik fontos könyvet szentelt neki, az állatok és növények variációját (1868)., Meggyőződése, hogy az evolúció volt arról, hogy a kis eltérések az a kis különbség, hogy az alkalmazkodás szervezetek, hogy életkörülményeik, ő csatolt döntő fontosságú, hogy kis mennyiségi eltérések, amelyek a rendszer utódai (különösen Karl Pearson), hogy megtalálta a biometryScience az élet mérés. Nagyon tág értelemben utal az élőlények mennyiségi tanulmányozására.. De Mendel törvényeinek 1900-as újrafelfedezése az érdeklődést a folytonos variációk felé fordította.
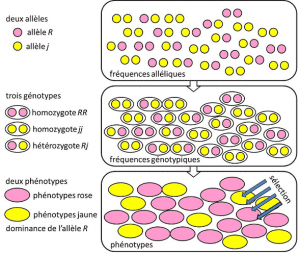
1908-Ból, hogy 1930-ban, a népesség genetikai alapvetően egy elméleti fegyelmet igyekszik összeegyeztetni Darwinizmus a Mendeli heredityHeredity alapján az átviteli egyetlen gén alatt egy domináns, recesszív, vagy nemi kromoszómához mód X (Y). Egy genetikai tulajdonságra utal, egyszerű determinizmussal, párral vagy kis számú génpárral., ahol a valószínűségek fontos szerepet játszottak. A genetika ellentétes intuitív., Azt jósolja, hogy leszármazottja nem reprodukálja a szülőt, mert az utóbbi genotípusa az allélok szétválasztásának lottójából származik, mielőtt a két szülői félgenom összeolvadna. Felismerték, hogy ezek az allélikus frekvenciákgyakoriság, amelynél egy változat allélja megtalálható egy populációban. Arányban vagy százalékban kifejezve. A populációban egy gén összes alléljának allélfrekvenciájának összege tehát definíció szerint 1. A populációgenetikában az allélfrekvencia a populáció vagy a faj szintjén a genetikai sokféleséget képviseli., generációk között terjednek, nem genotípusok vagy fenotípusok. Ezek a frekvenciák többé-kevésbé stabilak egyik generációról a másikra, és az egyének közötti filiációk többszörös összefonódásán túl ugyanazt a genotípusos gyakoriságot eredményezik. Az allél frekvenciák alapján határozva (4.ábra). A populációs genotípusok eloszlása, a varianciával összefüggésben, ezért az egyetlen kiszámítható tényező., 1930 körül a három teoretikus, Ronald Fisher, JBS Haldane és Sewall Wright segített megteremteni a Mendeliai népesség fogalmát, amelynek öröklődése engedelmeskedett Mendel törvényeinek. . Ebben az összefüggésben az evolúció szükségszerűen három szerkezeti szintet társít: a gént, az egyént és a populációt. Ez a háromutas társulás a következő képlettel foglalható össze: a populáció evolúciója az allélfrekvencia változása (gén skála, de csoport skála is), olyan változás, amely a fenotípusok szelekció szerinti válogatásától függ (egyedi skála)., Empirikus populáció genetika kutatási akkor fejlett, de korlátozott volt, hosszú ideje a tudatlanság, hogy kromoszómák munka (a DNS szerkezetét volt, megértette, 1953-ban, valamint a sorozatot kezdte, 1977-ben), majd vissza kellett vonulnom a tanulmány a pár látható polimorfizmusok, mint például a szín, a katicabogár elytra vagy a shell csiga (lásd a Hangsúly A nagy csiga vita). A genetikusok között sok vita volt arról, hogy a természetes populációk általában polimorfak vagy monomorfak-e, és a polimorfizmus önmagában előnyös-e vagy sem., A makromolekuláris skála variációjának vizsgálatát 1966-ig (a fehérjék esetében) és 1983-ig (a DNS esetében) meg kellett várni, mielőtt ezekre a kérdésekre válaszolnának.
kvantitatív variációk és örökölhetőség
amikor körülnézünk körülöttünk, fizikai különbségeket látunk a lények között, néhány összetett, mint az arcvonások, mások könnyen mérhetők, mint a súly vagy a magasság. A józan ész azt sugallja, hogy részben átvihetők,bár nehéz meghatározni. Ezek a tulajdonságok gyakran poligéneksok gén függősége alatt. A poligén öröklődésről beszélünk. A cukorbetegség poligén betegség.,, ami azt jelenti, hogy sok hely befolyásolja őket. A fejlődési genetika, valamint a fajok összehasonlítására való alkalmazása (evo-devo, a “fejlődés evolúciója”) feltárja a komplex kölcsönhatások hálóját, amelyek a fejlődés során a test sejtjei, mind azonos genetikai poggyászokkal, megkülönböztetik magukat epigenézissel, hogy különböző szöveteket adjanak. Még mindig nem világos, hogy összetett karakterek épülnek. A mérhető fenotípusokat biometrikus módszerekkel lehet tanulmányozni, a statisztikai eloszlások tulajdonságai alapján., Az egyik hasznos tulajdonságú érték a változásegy olyan mintában, amelyben egy adott tulajdonságot mértek, a variancia az értékek négyzetének átlaga és az értékek átlagának négyzete közötti különbség. Ez az intézkedés, amely mindig pozitív, az egyének szétszóródását jelzi. . Ha egy vonal variációját több Független ok határozza meg, akkor azok szórása additív, összegük pedig a vonal varianciáját adja. Ha nem függetlenek, akkor a kovariánsok összege hozzáadódik a varianciák összegéhez., A teljes variancia Vt a fenotípusos tulajdonság olyan, hogy :
Vt = Vga+Vgd+Vgd+Vgi+Ve
Vga, additív genetikai variancia, az az összeg, a szórások miatt minden locus függetlenül a többiek; Vgd, domináns genetikai variancia eredményei a kölcsönhatás a két allélek azonos locus; Vgi, kölcsönhatás eltérés között locuses, eredmények a tény, hogy a locuses ugyanannak a személynek befolyásolja; Ve, variancia, a környezet, feltételezzük, hogy itt a független genetikai variancia.,
a dominancia genetikai varianciája közös ugyanazon szülők két gyermeke számára, mert dominanciahatásaik vannak, amikor egy adott lokusban ugyanazokat az allélokat kapták mindkét szülőtől, ami velük történik minden negyedik locus. Nem osztják meg ezeket a hatásokat a szüleikkel, így inkább “hasonlóak” egymáshoz, mint a szüleikhez, annak ellenére, hogy oly sok gént osztanak meg egymással. Természetesen a szülő véletlenül ugyanazt a két allélt kaphatja, mint az egyik gyermeke egy adott helyen., Ez az esély a népesség alléljainak gyakoriságától függ. Látjuk, hogy a hasonlóság mértéke a gyermek és a szülő egy adott tulajdonság egy köztulajdon.
általánosabban, a genetikai variancia (VGA+Vgd+Vgi) minden összetevője populációk között változhat. A tenyésztési vonalakban gyakori eset a beltenyésztés, amely kimeríti a genetikai varianciát: ennek eredményeként egy tulajdonságnak ugyanaz az átlaga lehet két populációban, de eltérő genetikai variációval rendelkezik. A tenyésztő azon képessége, hogy kiválasszon egy tulajdonságot, ezért a vonal jellemzője, nem pedig a tulajdonság.,
a szülők és a gyermekek közötti kapcsolatot a H2 örökölhetőségével mérik. A meghatározás szerint az additív genetikai variancia aránya a teljes varianciához:
h2 = Vga/Vt

(2) korreláció 0, ahol a gyermekek mérete független, hogy a szülők. Egy értékes átlagos szülő közömbös értékű gyermeket hozott létre a gyermekek elosztásán belül. A pontok koordinátái egy körben helyezkednek el; értékük elvárása a szülők tengelyével párhuzamos vonalon (d) található., (3) közbenső szülő-gyermek korreláció 0 és 1 között: a pontok koordinátái ellipszisen helyezkednek el. Értékük várakozása már nem a koordinátatengelyek felezővonalán található( amely szintén az eloszlás ellipszis fő tengelye), hanem az r regressziós vonalon. egy átlagos érték szülője b értékű gyermeket hozott létre (0 és a között) egy bizonyos varianciával. Megjegyzendő, hogy a (3) eset köztes az (1) és (2)
esetek között, az 5.ábrán különböző feltételezések szerint., Az örökölhetőség az a képesség, hogy meghatározzuk, hogy kiválaszthatunk-e egy funkciót. 5-3 ábra azt mutatja, hogy egy tenyésztő lesz képes belül átkelés generációs, hogy mozog az átlagos, egy karakter a 0 értéket az érték, b, ha kiválasztja azt az egyének értéke egy. Meg tudjuk mutatni, hogy a b = h2a. Válassza ki a tulajdonság hasznos, hogy növelje a heritability arány által standardizing a környezet (így csökkentve a környezeti variancia), valamint azáltal, hogy magát a környezeti feltételek, amelyek mellett a megjelenése a tulajdonság., Waddington genetikus kimutatta, hogy a természetben a környezet változása így új tulajdonságokat fedhet fel, amelyeket az evolúció során választanak ki.
referenciák és megjegyzések
borítókép. Csigahéjak Cepaea nemoralis & Cepaea hortensis. André Künzelmann, UFZ.,
Ez a képlet hagyományosabb módon írható: \pi = \ frac{n (n-1))} {2L}\sum _{I=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
olyan egyének mintája, akiknél a vizsgált tulajdonságot mértük, a variancia meghatározható az értékek négyzetének átlaga és az értékek átlagának négyzete közötti különbségnek. Ez az intézkedés, amely mindig pozitív, az egyének szétszóródását jelzi.
Ez a kifejezés szigorú értelemben vett örökölhetősége. Az örökölhetőség tág értelemben a genetikai varianciák összegének a teljes varianciához viszonyított aránya., Ez a képlet adja örökölhetőség mindkét szülő (az “átlagos szülő”); a becslés csak egy szülő adna h2/2.
a környezet környezeti enciklopédiája az Association des Encyclopédies de l ‘environnement et de l’ Energie (www.a3e.fr), a grenoble-i és a Grenoble-i Egyetemhez, valamint a Francia Tudományos Akadémia támogatásával.
a cikk idézéséhez: VEUILLE Michel (2021), genetikai polimorfizmus és variáció, enciklopédia a környezetről, url : https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
A cikkek Enciklopédia, a Környezet rendelkezésre értelmében a Creative Commons BY-NC-SA licenc, amely felhatalmazza a szaporodás feltétele: hivatkozva a forrás, nem kereskedelmi célú használata őket, megosztása azonos kezdeti feltételek, lejátszó minden újrafelhasználás vagy elosztása a megemlíti, hogy ez a Creative Commons BY-NC-SA licenc.
Vélemény, hozzászólás?