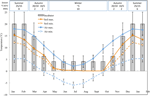
1.ábra. Havi maximális és minimális levegő – és talajhőmérsékleti adatok az Ausztrál Alpokban (átlag ± STDEV). A levegő hőmérséklete a BOM adatbázis átlaga, a talaj hőmérsékletét ibuttonokkal mértük (lásd a módszereket). A rudak a kísérleti inkubátor hőmérsékletét jelzik (átlag ± STDEV)., Az évszaknak megfelelő idő (évszak), inkubátor hőmérséklet-szabályozás (°C, 12/12 h, világos/sötét) és a kezelés időtartama (hetek) a felső panelen feltüntetett módon.
Azzal a céllal, hogy feltárása hőmérséklet közel, hogy azok a magok élmény utáni szétszóródását, a napi maximális, illetve minimális a talaj hőmérséklete az adatgyűjtés belül a tanulmány helyszínen (Tengerész Kunyhó; 36.27°S 148.17°E, 2030 m.ok.l.) használata ibutton adatok favágók (n = 10, Beágyazott Adatok a Rendszerek, USA) elhelyezett 4 cm-rel a talaj felszínén a bázison a növényzet január 17-től December 17-ig 2012., A felszíni zavar elkerülése érdekében 4 cm-re helyeztük el a favágókat, de továbbra is reprezentatívak azok a feltételek, amelyeknek a magok a talajban vannak kitéve; ez a mélység köztes az alpesi talajhőmérséklet-ellenőrző gyakorlatokban (Scherrer and Körner, 2011; Pauli et al., 2015). A gyanú szerint az átlagos talajhőmérséklet télen nem esett fagypont alá (1. ábra). Nyáron a növényzet alatti talajhőmérsékletről ismert, hogy nyomon követi a környezeti hőmérsékletet, míg a csupasz talaj hőmérséklete legfeljebb 15°C–kal haladja meg a környezeti hőmérsékletet (a talajvédelmi szolgálat nem publikált nyilvántartása, 1960-as évek-1970-es évek).,
16 családból és 37 nemzetségből származó 54 faj érett magjait gyűjtötték össze 2009.január és 2010. április, valamint 2011. január és április között (a teljes neveket és hatóságokat lásd az 1. táblázatban). Utalványokat nyújtottak be az Ausztrál Nemzeti herbárium, Canberra). Összességében a fajok az alpesi régiókban található Ausztrál angiosperm flóra több mint egynegyedét képviselték (Costin et al., 2000, lásd az 1. táblázatot), bár ezek közül a fajok közül sok a treeline alatt is kiterjed., Az összes gyűjtemény életképességét a vetés előtt becsülték meg kísérleti csírázási körülmények között a tetrazolium-klorid (TZ) festési technikával (International Seed Testing Association, 2003). A gyűjtéssel és feldolgozással kapcsolatos további részletekért lásd a Kiegészítő anyag A. függelékét.
1. táblázat
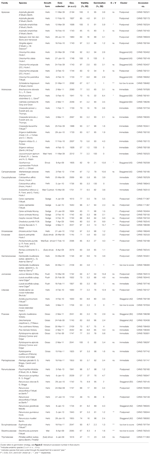
1.táblázat. A vizsgálati fajok, a család és a hatóság, a növekedési forma és a begyűjtési adatok, a vetőmag százalékos életképessége (TZ-becsült, ± S. E.), Az elért végső százalékos csírázás (±S. E.,), az életképesség és a csírázás közötti statisztikai különbség (g < V, amelyet csak olyan esetekben vizsgáltak, amikor a csírázás kisebb volt, mint az életképesség).
1.táblázat. A vizsgálati fajok, a család és a hatóság, a növekedési forma és a begyűjtési adatok, a vetőmag százalékos életképessége (TZ-becsült, ± S. E.), Az elért végső százalékos csírázás (±S. E.,), az életképesség és a csírázás közötti statisztikai különbség (g < V, amelyet csak olyan esetekben vizsgáltak, amikor a csírázás kisebb volt, mint az életképesség).
Phenology a csírázási vizsgálta utánozva hőmérséklet rendszerek alpesi magok tapasztalat in situ, post-szétszóródását, egy mesterségesen rövidített progresszió a szezont. A vetőmagokat a kísérlet során átitatták, így az eredmények csírázási potenciált jeleznek, ha a víz nem korlátozza., Levegő, talaj, hőmérséklet adatokat (1. Ábra) használtak útmutató, majd érvényesítse inkubátor hőmérséklet-rendszerek, bár a logisztikai contraints dolgozom inkubátorok kizárt beépítése az ingadozás jellemző, hogy természetes körülmények között. A csírázási tesztek gyűjteményenként nyolc, 25 magból álló replikációt használtak, amelyeket 9 cm átmérőjű műanyag Petri edényekbe vetettek, amelyek 1% sima víz-agart tartalmaznak. A Petri-edényeket Parafilm segítségével lezárták, hogy elkerüljék az agar kiszáradását, mielőtt csírázási inkubátorokba helyezték (Thermoline Scientific, Melbourne, NSW, Ausztrália)., Az edények felét alumíniumfóliába csomagolták, hogy kizárják a fényt, és jelezzék a csírázásra vonatkozó fénykövetelményeket. A gyűjtésenkénti fénykezelésenkénti négy replikáció mindegyikét egy másik polcra helyezték ugyanazon inkubátoron belül, és a polcon lévő helyet hetente újrarendezték. Minden ismétlések minden gyűjtemény volt költözött át a következő szimulált thermoperiods: 8 hét 25/15°C-ig (nap/éjszakai hőmérséklet) → 2 hét 20/10°C → 2 hét 10/5°C → 10 hetes állandó 4-5°C → 2 hét 10/5°C → 2 hét 20/10°C → 6 hét 25/15°C., Így az egész kísérlet egy ciklusa utánozta a nyárra, kora őszre, késő őszre, télre, kora tavaszra, késő tavaszra és nyárra emlékeztető hőmérsékleteket, amelyek összesen 32 hétig tartottak. A rövidített évet ésszerűnek tekintették, mivel a csatlakozások általában egy adott kezelés során megszüntették a további csírázást, mielőtt a következő “szezonra”váltottak volna., A fénycsövek (> 90 µmol m−2 s−1 minden polc közepén) 12/12 órás fény/sötét fotoperiódust biztosítottak, a tavaszi, nyári és őszi hőmérsékleti viszonyok pedig egybeestek a nap meleg hőmérsékleti időszakával. A több mint 1 mm-es radicle-megjelenésként definiált csírázást 7 naponta pontozták, majd a csírázott magokat és palántákat később eltávolították az edényekből. A fóliába csomagolt lemezek csírázását a fentiek szerint értékelték, de nagyon alacsony zöld fény mellett.,
egy kísérleti ciklust (32 hét) követően a legtöbb gyűjtemény megszűnt (lásd a kivételek kiegészítő adatait), miután ebben az időben legalább 2 hétig nulla csírázást mutatott. A befejezést követően az összes megmaradt ép magot szikével boncolták mikroszkóp alatt. A szilárd, friss endospermiummal és embrióval rendelkező vetőmagokat életképesnek tekintették, és az embrióból kiürült magokat levonták a teljes összegből az egyes gyűjtemények százalékos csírázásának kiszámításakor., Kiszámoltuk, egy index a fény követelmények csírázási az egyes fajok, mint egy töredéke a csírázás, a sötét kezelés abban az időben, hogy kumulatív csírázás százalékosztályok 50 95% – a elérte a váltakozó fény/sötét kezelés (lásd alább).
A Csírázási stratégia ökológiai korrelációi
összeállítottunk egy adatbázist a fajok funkcionális tulajdonságairól és földrajzi intézkedéseiről a közzétett adatokból a csírázási stratégiával való korreláció értékelésére., Az adatbázis a következő folytonos jellemzőket tartalmazta: átlagos magtömeg, a csírázásra vonatkozó könnyű követelmény (lásd fent), fajlagos levélterület (friss terület/száraz tömeg), egyedi levélterület, egyedi levélszárazság, valamint Érettségi magasság. A faj minimális, átlagos és maximális begyűjtési emelkedését az Ausztrál virtuális herbárium összes herbárium-nyilvántartása alapján határozták meg, és a vizsgálatban használt speciális vetőmaggyűjtemények magasságát is folyamatos tulajdonságként vették figyelembe. A növekedési formát három kategóriába sorolták: graminoid, gyógynövény vagy cserje., Egy bináris tulajdonságot is tartalmaztunk: függetlenül attól, hogy a magok endospermikusak-e vagy sem.
a tulajdonságadatokat számos forrásból gyűjtötték, köztük az Ausztrál virtuális Herbáriumból (http://avh.chah.org.au/), az Ausztrál Nemzeti herbárium mintainformációs nyilvántartásból (https://www.anbg.gov.au/cgi-bin/anhsir), a Kosciuszko alpesi flórából (Costin et al., 2000), az Új-Dél-Wales-i Flóra (Harden, 1990) és a magok összehasonlító belső morfológiája (Martin, 1946). A vetőmag tömegét és a levél specifikus területét a csírázási kísérletek során használt egyes mezőgyűjteményekből nyerték., A vetőmag tömegét négy tétel 10 mag tömegének mérésével nyerték. Az SLA-t öt, fajonként különálló egyedekből származó, teljesen kiterjesztett levél alapján határozták meg. Ezek voltak a terepen gyűjtött, szkennelt a síkágyas szkenner, szárított 60°C-on tömegállandóságig súlyozott átlagával, hogy a legközelebbi 0.000 g. Ahol elegendő vetőmag állt rendelkezésre, vagy a specifikus levélterület nem állt rendelkezésre adatok származnak, a többi ANBG gyűjtemények ugyanaz a faj, vagy más adatforrásból (S. Venn, nem publikált adatok)., A vetőmag endospermikus volt-e vagy sem az ANGB seed bank nyilvántartásában szereplő vetőmag rajzai alapján határozták meg az ANBG seed bank munkatársaival konzultálva. A teljes adatbázis és források kérésre a szerzők.
Adatelemzés
a csírázási mintákat vizuálisan értékelték, és a PC-ORD for Windows klaszterelemzésével, a 6.verzióval (Mjm Software Design, Gleneden Beach, vagy, USA). Az analízis során euklideszi távolságmátrixot és egy legközelebbi szomszéd kötési algoritmust alkalmaztak., A klaszterelemzés adatpontjai az egyes gyűjtéseknél (T20, t40, t50, t60, t80 és t95%) a specifikus kumulatív csírázási százalékokhoz viszonyított idő (hetek) voltak (átlagosan a 4 edényen). Így egy gyűjtemény, amely kicsírázott elején a kísérlet volna rövid alkalommal százalékosztályok akár t80% például, mivel a gyűjtemény, hogy elhalasztották a csírázás, amíg a hőmérséklet emelkedett következő a szimulált téli volna régen, hogy ezek a százalékosztályok., Összetettebb csírázási minták, például lassú, egyenletes csírázás vagy csírázási kitörések is kimutathatók, amelyeket a kísérlet kezdetén és végén egyenletesen elosztott időpontok vagy időszakok jeleznek. Ezt a módszert alkalmazták a csírázási minta objektív értékelésének előírására, függetlenül a hőmérséklettől.,
egyirányú varianciaanalízis (ANOVA) végezték a párhuzamos lemezek minden gyűjtemény, hogy felmérje, vajon van-e szignifikáns különbség százalékos életképesség (által meghatározott TZ-teszt), illetve a végső százalékos csírázás. További általános lineáris modelleket alkalmaztak annak értékelésére, hogy a klaszterelemzésből származó klaszterek életképességükben vagy végső csírázási százalékukban különböznek-e.
az ökológiai vonások és a csírázási stratégia klaszterek közötti társulásokat kétféle módon tesztelték., Először a klaszterek közötti átlagos különbségeket teszteltük, az ANOVA-t a klaszterrel rögzített hatásként, az ökológiai korrelációkat pedig a válaszváltozókként, vagy az endospermia és a növekedési forma replikált G-tesztjeit használva. Másodszor, mivel a csírázást és más funkcionális tulajdonságokat nagy valószínűséggel befolyásolja az evolúciós történelem, ezeket a korrelációkat filogenetikai regressziók alkalmazásával értékeltük. A phylogenyt a phylocom Phylomatic és a bladj packing in R (Webb et al., 2008; R Core Team, 2013)., A filogenetikai szerkezet jelenlétét a változókban a phylosig vagy a phylo segítségével mértük fel.d (diszkrét vagy bináris változók) a Phytools csomag funkciója R-ben (Revell, 2012). A filogenetikai regressziókat a caper (Orme et al., 2013). Az elemzéseket strukturált módon végezték, tükrözve a PC-Ord-ben feltárt klasztert (lásd alább)., Kezdetben négy csírázási stratégiát (elhalasztott, azonnali és lépcsőzetes kategóriákat az előzőekben), majd három (az összes lépcsőzetes faj összeillesztése és a halasztott és közbenső), végül pedig a két legszélesebb stratégia, a halasztott és a közbenső közötti különbségeket vizsgáltuk.
eredmények
A vetőmag életképessége a végső csírázáshoz képest
összességében a vetőmag életképessége és a végső csírázási arány magas volt. Átlagos TZ-becsült életképesség az összes gyűjteményben 80 ± 1 volt.,7%, a gyűjtemények több mint kétharmada több mint 75% – os életképességet mutat (1. táblázat). A gyűjtemények minőségében nem volt különbség a 3 év alatt (az átlagos életképesség 2009-ben, 2010-ben és 2011-ben 79 ± 2,8, 80 ± 2,7, illetve 81 ± 3,7% volt), ami azt jelzi, hogy a 2009-ben gyűjtött vetőmagok bankja nem csökkentette életképességüket.
Az 54 fajból 45 esetében sikerült a tz-nél nagyobb vagy nem szignifikánsan alacsonyabb csírázást elérni (egyirányú ANOVA: p> 0, 05, 1.táblázat)., A fennmaradó kilenc faj közül négy faj csírázása nagyon alacsony volt (< 5%), a jó életképesség ellenére (kb. 55-94%), így nem tudtunk következtetéseket levonni e fajok csírázási stratégiáiról. A csírázás elmulasztása azt tükrözheti, hogy kezeléseink nem enyhítették a nyugalmat, vagy hogy a magok nem voltak teljesen érettek a gyűjteményben. A kísérlet végén végzett vágásvizsgálatok azt mutatták, hogy minden esetben a nem csírázott magok többsége egészséges, imbibed embriót tartalmazott., Érdekes módon 34 faj olyan csírázást ért el, amely nagyobb volt, mint a tz becsült életképessége (1.táblázat), ami arra utal, hogy a tz-teszt általában alábecsülte a vetőmag életképességét.
Csírázási klaszterek
klaszterelemzés két fő klasztert (A és B) tárt fel, mindegyikben alszerkezettel (2.ábra). A fajok többsége (n = 21 faj 10 családból) esetében a csírázást elhalasztották a téli hőmérséklet (5°C nappali/éjszakai) időtartamára, vagy amíg a hőmérsékletet tél után 10/5°C-ra emelték (A1 és A1:tél, 2., 3a. ábra)., Alternatív megoldásként a csírázás azonnal 25/15°C-on kezdődött (n = 17 faj, hat család, a B1 klaszterben, 2.ábra, 3b). E fajok többsége esetében a csírázási arány gyors volt; a teljes csírázás (t50) 50% – a kevesebb, mint 3 hét alatt történt (1.Kiegészítő ábra).
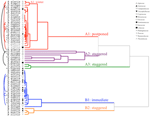
2.ábra. Klaszterelemzés dendrogram a kumulatív csírázási százalékig eltelt idő alapján(20, 40, 50, 60, 80, és 95%). Faj, amelyet a faj eredeti és első része jelöl. A csillagok olyan gyűjteményeket jelölnek, amelyeket a tesztelés előtt tiltottak., A finom ívek összekapcsolják a duplikált gyűjteményeket, hogy összehasonlítsák a banki tevékenység csírázásra gyakorolt hatását. A szimbólumok különböző családokat jelölnek (lásd a legendát). A szürke szövegben bemutatott gyűjtemények azok, amelyek a kísérlet két ciklusának voltak kitéve.
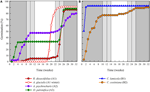
3.ábra. Az egyes csírázási klaszterekből egy reprezentatív faj kumulatív százalékos csírázása. A) A klaszter, a csírázást elhalasztó Fajok, B) B klaszter, azonnal csírázó Fajok., Vegye figyelembe,hogy a lépcsőzetes csírázási mintákat a fenti klaszterekbe ágyazzák. Az inkubátor hőmérsékleti rendszereit árnyékolás képviseli (lásd az 1.ábrát).
Ezen széles osztályok mindegyikén belül azonban egyértelmű alcsoportok voltak, amelyek tükrözik a csírázást, amely az idő múlásával eltolódott, azaz a vetőmag-tétel egy része a téli hőmérsékletnek való kitettség előtt csírázott, míg a fennmaradó rész a tél után csírázott. A fokozatos csírázás hat család 13 fajában történt,és elsősorban akkor, amikor a nappali hőmérséklet meghaladta a 10°C-ot (3a., B. ábra)., Ezeket a lépcsőzetes csírázókat három csoportra osztották a klaszterelemzésben. A hét gyűjteményű A2 klaszter jelentős csírázást mutatott mind a hideg hőmérsékleti időszak előtt, mind azt követően. Ahol a csírázás a hőmérséklet megváltoztatása előtt megszűnt, arra a következtetésre jutottunk, hogy a tél előtti további csírázás hiányát nem a hőmérséklet (A3) okozta. Ezzel szemben az A2-csoport nagy részének csírázása csak akkor állt meg, ha a hőmérsékletet 10/5°C-ra csökkentették., A B2 klaszter további négy fajt tartalmazott, amelyek a kísérlet korai szakaszában jelentős csírázást mutattak, és a teljes csírázásuk viszonylag kis részét elérték, miután a hőmérsékletet 5°C fölé tettük (3b ábra).
Az A1 és B1 klaszterekben a fajok közötti viszonylag kisebb eltéréshez képest a szakaszos klaszterek sokkal nagyobb különbséget mutatnak a fajok között. Az egyes görbék minden fajra vonatkozóan az 1i–vii. kiegészítő ábrán láthatók.a csírázási stratégiával kapcsolatos további részletekért lásd a Kiegészítő anyag B. függelékét.,
Ökológiai Összefügg a Csírázás Stratégia
az ökológiai összefügg, hogy mi tekinthető, csak a közvetlenül kapcsolódó mag tulajdonságok szignifikánsan korrelál a csírázás stratégia, függetlenül attól, hogy tudjuk értékelni a korrelációs szinten két nagyobb klaszterek (elhalasztott vs azonnali csírázás), vagy szintjén három, vagy négy klaszterek, vagy akár elszámolni filogenetikai szerkezet vagy nem (2.Táblázat)., Az elhalasztott csírázással rendelkező fajok nehezebb magvakkal rendelkeztek, és nagyobb valószínűséggel voltak endospermikusak, mint az azonnal csírázó Fajok (4.ábra). A szakaszos csírázással rendelkező fajok a vetőmag tömegében és az endospermiában köztes jellegűek voltak. Azonban nem minden magjellemző korrelált a csírázási stratégiával: nem volt összefüggés a vetőmag csírázására és a stratégiára vonatkozó könnyű követelmények között. Várakozásainkkal ellentétben a magasabb fajlagos levélterületű fajok (amelyek potenciálisan magasabb növekedési rátát jeleznek) nem voltak nagyobb valószínűséggel azonnali csírázást mutatnak., Hasonlóképpen, a magasabb átlagos magasságú vagy kisebb magassági tartományú fajok nem voltak nagyobb valószínűséggel elhalasztott csírázást mutatnak.

2. táblázat. A filogenetikai szerkezet előfordulása a csírázási stratégiában és annak összefüggésében.
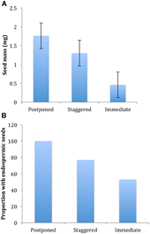
4. ábra., Az a) Átlagos vetőmagtömegben és B) az endospermikus magvakkal rendelkező fajok aránya az elhalasztott (A1 klaszter), lépcsőzetes (A2, A3 és B2 klaszter kombinációja) és azonnali csírázási (B1 klaszter) stratégiák között. A stratégiák közötti különbségek jelentősek a p ≤ 0.05 értéknél, lásd a 2.táblázatot.
a Vita
Csírázási az alpesi élőhelyek volt történelmileg úgy kell tekinteni, hogy a környezetvédelmi szempontból ellenőrzött, a téli hó szigetelő mag ellen potenciális csírázás jeleket, visszaadás nyugalmi felesleges (Billings, valamint Mooney, 1968)., Eredményeink azonban alátámasztják a legutóbbi bizonyítékokat arról, hogy a nyugalmi mechanizmusok gyakoriak az alpesi Fajok között mind Ausztráliában (Hoyle et al., 2013; Sommerville et al., 2013) és máshol a világon (Cavieres and Arroyo, 2000; Schutz, 2002; Shimono and Kudo, 2005; Mondoni et al., 2009; Schwienbacher et al., 2011)., Nyugalmi tűnhet, hogy ez jelentős szerepet fog játszani irányítja az in situ időzítés a csírázás sok Ausztrál alpesi faj eljárva, hogy késedelem, elhalasztása, vagy lassú az arány a csírázás, így potenciálisan biztosító kockázatkerülő regeneráció stratégiák a kemény, változó környezetben. Csírázási eredményeink közül talán a legszembetűnőbb azon fajok aránya volt, amelyek csírázási stratégiája egy adott faj vetőmaggyűjteményében változott, ami arra utal, hogy mind a nyugvó, mind a nem alvó tulajdonságokat ugyanazon faj vetőmaggyűjteményében mutatták ki., Nevezetesen, a csírázási stratégiák általában erősen konzerváltak a családokon belül, és korreláltak a vetőmag anatómiájának elemeivel: a tömeggel és az endospermiával. A csírázási stratégiák azonban függetlenek voltak az érett növény Egyéb ökológiailag jelentős funkcionális tulajdonságaitól.
A Csírázási stratégia változása
a vizsgált fajok közel felének magjai nem tudtak csírázni, amíg hűvös, nedves időszaknak (állandó 5°C) nem voltak kitéve, ami arra utal, hogy a hideg rétegződés enyhítette a fiziológiai nyugalmi mechanizmust., A csírázás elhalasztása a következő tavaszig lehetővé teheti a palánták számára, hogy elkerüljék a kemény tél felett vagy előtt történő letelepedést, miközben optimalizálják a következő növekedési időszakot is. Ezzel szemben az azonnal csírázó fajok által mutatott opportunista csírázás szelektív előnyt jelenthet, ha a téli palánták elhullásának kockázata alacsony, lehetővé téve a növények virágzását a következő tavasszal vagy nagyobb méretben (Donohue, 2002).,
eddig kevés közzétett bizonyíték volt az alpesi vetőmaggyűjteményekben a nyugalmi mechanizmusok intraspecifikus változékonyságára, mint például az itt megfigyelt lépcsőzetes csírázási stratégiák magyarázata. A csírázás lépcsőzetes az idő múlásával is magyarázható különböző szintű mag lejárat egyének között a lakosság a gyűjtés időpontjában, és/vagy jelzi, hogy dimorf stratégia belül egyetlen növény kapcsolódó helyzetben, a növény vagy az időzítés a fejlődés.,
Az evolúciós tét-fedezeti ügyleteket gyakran arra késztetik, hogy megmagyarázzák a vetőmag csírázási jellemzőinek diverzifikált stratégiáját (Simons, 2011), például ezt a szakaszos csírázással rendelkező gyűjtemények bizonyítják. Ha azonban egy diverzifikált stratégia mind az átlagos fitnesz növekedését, mind a variancia csökkenését eredményezi (például a testvérek közötti verseny csökkentésével), akkor ez nem minősül bet-fedezeti ügyletnek. Hasonlóképpen, ha a látszólagos diverzifikáció az egyének közötti különbségeket tükrözi, nem belül, akkor nem tétfedezés (Starrfelt and Kokko, 2012)., Vizsgálatot alpesi csemete terjedési mintázat, valamint a tolerancia, a fagy, a hó olvad, párosítva vizsgálatok a befolyásoló tényezők a csírázás stratégia variáció faj, megmagyarázza a csírázás phenology a fajok kialakulása lépcsőzetesen stratégia, valamint jelzi, hogy valószínűleg, hogy képviselje a tét-fedezeti stratégiák.,
A Csírázási stratégia ökológiai korrelációi
a csírázási stratégia és más ökológiailag fontos tulajdonságok közötti kapcsolat vizsgálata kimutatta, hogy a csírázási tulajdonságok korreláltak más magjellemzőkkel, de nem a vegetatív tulajdonságokkal. Sommerville et al. (2013) megállapította, hogy a nem endospermikus magvakkal rendelkező Ausztrál alpesi gyógynövények nagyobb valószínűséggel nem szunnyadnak a szétszóródáskor. Munkánk alátámasztja ezt a megállapítást, továbbá azt is jelzi, hogy a kisebb magok nagyobb valószínűséggel lesznek a közvetlen csírázók között. Bár Sommerville et al., (2013) nem közölte, hogy az endospermia miért társulhat a nyugalmi állapothoz, azt javasoljuk, hogy a tartalékok sziklevelekben történő elhelyezése javíthatja a korai növekedési rátát és a közvetlen csírázó fajokban történő letelepedést. Eredményeink nem mutattak összefüggést a Fényigény és a nyugalmi állapot között, bár a Fényigény fajonként eltérő volt. Számos más magjellemző is létezik, amelyek változó környezetben pufferelik a kihalás kockázatát, például a vetőmag-diszpergáló mechanizmusok és a vetőmag-élettartam a talajmagban (lásd Tielborger et al., 2012 felülvizsgálatra)., Ebben a szakaszban nem tudjuk megmondani, hogy ezek a tulajdonságok korrelációt mutatnak-e a csírázási stratégiával, vagy sem.
azt is megkérdeztük, hogy a csírázási stratégia összefügg-e az ökológiailag fontos nem reproduktív tulajdonságokkal, például a levélterület vagy a növény magassága (Westoby, 1998; Wright et al., 2004). Különösen azt állítottuk, hogy az azonnali csírázó fajok olyan tulajdonságértékeket mutathatnak, amelyek a gyorsabb növekedési rátát jelzik (az SLA-t proxyként használva), ami a szétszóródás és a tél kezdete közötti rövid időszakban szükséges lehet., Azonban nem találtunk ilyen összefüggéseket. Érdemes megfontolni, hogy a palánták élettani és növekedési tulajdonságai a korai letelepedési szakaszban eltérő mintákat mutathatnak-e, mint a felnőtt vegetatív tulajdonságok, de a jelenlegi munka hatókörén túl.
végül megvizsgáltuk, hogy a magasabb magasságokból származó fajok, ahol a vegetációs évszakok általában rövidebbek, nagyobb valószínűséggel mutatnak-e nyugalmi állapotot, de nem volt összefüggés sem a magasságtartomány, sem a gyűjtési magasság, sem a gyűjtemény egészének átlagos emelkedése és a csírázási stratégia között., Ezek az eredmények együttesen azt mutatják, hogy míg a csírázási stratégia fejlődése szorosan kapcsolódik az ökológiailag fontos vetőmagjellemzőkhöz, egy nagyrészt független tengelyt tartalmaz a vegetatív vagy elosztási jellemzőkhöz.
hatással Alpesi Növény Közösségek a Változó Éghajlat
Ausztrál alpesi ökoszisztémák, mint az alpesi területeken szerte a világon veszély fenyegeti a klímaváltozás együtt változik a tűz frekvencia, földhasználat minták, beáramlása az invazív fajok, a hatások, a megnövekedett emberi látogatás., Az éghajlatváltozással kapcsolatos felmelegedés gyorsabban megy végbe a fák felett, mint az alacsonyabb magasságokban, és az alpesi területek várhatóan továbbra is az átlag feletti felmelegedést tapasztalják a jövőben (Kullman, 2004). Az alpesi növényeknek az éghajlatra és a kapcsolódó változásokra adott válaszában valószínűleg jelentős szerepet játszik az új körülmények között a vetőmagon keresztüli folyamatos regeneráció képessége. Emellett a megváltozott zavarrendszerek valószínűleg nagyobb szerepet igényelnek a régióban a helyreállításban és a rehabilitációban., A betekintést az itt közölt ezért fontos, hogy megőrzése, kezelése alpesi rendszerek alatt változás; segíthetnek tájékoztatja a jóslatok, hogy milyen vizsgálat faj hatással lehet képezik az alapját a vetőmag-szaporítás tervek ezek a fajok, illetve használható útmutató jövőben nyomozást alpesi csírázási stratégiák azonosítható conservation and management eredmények.,
a hőmérséklet változása valószínűleg befolyásolja a magok nyugalmi állapotát mind a pre -, mind a poszt-diszpergálódás során, következésképpen megváltoztatja és / vagy megzavarja a csírázási stratégiákat, ami a jelenlegi csírázási fenológia eltolódását eredményezi (Hoyle et al., 2008b). Az alpesi növények esetében az éghajlat felmelegedése tavasztól őszig terjedő változáshoz vezethet, amelyet elsősorban a vetőmag nyugalmi állapotának változása okoz, és ami jelentős következményekkel jár a tavasszal megjelenő fajokra (Mondoni et al., 2012)., További kutatások hogyan változik a hőmérséklet, hogy a magok élmény utáni szétszóródását, csökkentett időtartama hótakaró, illetve gyakoribb a szélsőséges hőmérséklet események hatással lesz a csírázás érdekében szükséges megjósolni, hogy faj a nyugalmi válaszol, hogy az éghajlatváltozás. Azoknak az alpesi fajoknak, amelyek ősszel, a fagy és a havazás előtt csíráznak, eredményeink felvetik azt a kérdést, hogy ezeknek a fajoknak a palántái hogyan fognak megbirkózni a csapadék várható csökkenésével és az a-szezonális fagy események megnövekedett gyakoriságával.,
Végre, az eredmények emelni fontos kérdése variáció fajok, különösen a szövetség előrejelzése az éghajlatváltozás hatása, vagy kiválasztja, hogy melyik magokat, hogy kihasználja a helyreállítás. A csírázás stratégiák eltérőek lehetnek belül a faj, attól függően, lakosság, virágzási idő, mag tömege, szülői befektetés, éghajlat, domborzat, és/vagy anyai környezet, ez a változás tükrözheti genetikai, epigenetikai, és/vagy a környezeti tényezők (Cochrane et al., 2015)., Növekvő magasság számlák variáció csírázhatóság és / vagy nyugalmi állapotát több alpesi Fajok (Billings and Mooney, 1968; Dorne, 1981; Cavieres and Arroyo, 2000), de lásd (Hoyle et al., 2014). A csökkent nyugalmi állapot a környezeti feszültségekből eredő vetőmagfejlesztés korai megszüntetéséből eredhet (Steadman et al., 2006; Hoyle et al., 2008b)., Vizsgálata változása csírázás stratégiák belül faj is mutatják, hogy a potenciális fajok megbirkózni, valamint alkalmazkodni, az éghajlat megváltozása, így fontos szerepet játszik meghatározó jövőben, a túlélés, a fajok disztribúciók, amelyek fontos betekintést a kezelése, megőrzése.
következtetések
megközelítésünk feltárta az Ausztrál alpesi növényvilágban a nyugalmi állapot széles körű előfordulását, és bizonyítja, hogy a csírázási stratégiák a vetőmaggyűjteményen belül, valamint a gyűjtemények között változnak., Továbbá megmutattuk, hogy míg a nyugalmi stratégia filogenetikailag konzervált és korrelál a vetőmag tömegével és az endospermiával, ez nagyrészt független a vegetatív vonásoktól és a tartomány jellemzőitől. Bár nem lehet közvetlenül az eredményeinkből extrapolálni, hogy az éghajlat és a kapcsolódó változások hogyan befolyásolják az alpesi növényzetet, ezek az eredmények képezhetik a szaporítási tervek tervezésének alapját, valamint a változó éghajlati rendszerek hatására irányuló további kísérleteket., Különösen javasoljuk, hogy további kutatások megértése okai belül-fajok változása csírázás stratégia, valamint a szerepet játszhat abban, hogy képes a faj, hogy ellenálljon megjósolta, hogy az éghajlatváltozás.
Data Accessibility
a teljes csírázási adat – és tulajdonságadatbázis a DRYAD-on, valamint az élő Ausztrália atlaszában kerül elhelyezésre.
összeférhetetlenségi nyilatkozat
a szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.,
elismerjük az Ausztrál Kutatási Tanács (LP0991593) és az Ausztrál Nemzeti Botanikus kert és annak barátai támogatását. Köszönjük J. McAuliffe, S. Fethers és S. Lawatsch az ANBG. Köszönetet mondunk továbbá M. Holloway-Phillips, M. Cardillo és S. Cunningham-nek a kézirat korábbi változataihoz való hozzájárulásért, valamint analitikai tanácsadásért és S. Venn-nek a tulajdonságadatokhoz való hozzáférésért.
Kiegészítő anyag
Foley, M. E. (1992). Az oldható cukrok és gibberellinsav hatása a kimetszett zab (Avena fatua) embriók lebontására., Gyomirtó. 40, 208–214.
Google Scholar
Harden, G. J. (1990). Új-Dél-Wales növényvilága. Sydney, NSW: Royal Botanic Gardens.
Google Scholar
International Seed Testing Association. (2003). Munka lapok Tetrazolium vizsgálat. Bassersdorf: ISTA.
Kullman, L. (2004). Az alpesi világ változó arca. Glob. Változás Newsl. 57, 12–14.
Google Scholar
Vélemény, hozzászólás?