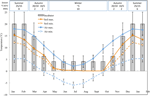
Figura 1. Dati mensili di temperatura massima e minima dell’aria e del suolo nelle Alpi australiane (media ± STDEV). Le temperature dell’aria sono medie dal database BOM, le temperature del suolo sono state misurate con iButtons (vedi metodi). Le barre indicano le temperature sperimentali dell’incubatore (media ± STDEV)., Il periodo equivalente dell’anno (stagione), il regime di temperatura dell’incubatore (°C, 12/12 h, chiaro/scuro) e la durata del trattamento (settimane) come indicato nel pannello superiore.
per scoprire le condizioni di temperatura vicina a coloro che semi esperienza post-dispersione, giornaliero minimo e massimo di temperatura del terreno sono stati raccolti i dati all’interno del sito di studio (da Marinaio Capanna; 36.27°S e 148.17°E, 2030 m.s.l.) utilizzando ibutton registratori di dati (n = 10, Embedded Sistemi di Dati, USA) posto 4 cm sotto la superficie del suolo alla base della vegetazione dal 17 gennaio al 17 dicembre 2012., Abbiamo posizionato i logger a 4 cm per evitare disturbi superficiali ma essere comunque rappresentativi delle condizioni a cui i semi sono esposti nel terreno; questa profondità è intermedia a quella utilizzata in precedenti esercizi di monitoraggio della temperatura del suolo alpino (Scherrer e Körner, 2011; Pauli et al., 2015). Come si sospettava, la temperatura media del suolo non è scesa sotto lo zero durante l’inverno (Figura 1). In estate, la temperatura del suolo sotto la vegetazione è nota per monitorare la temperatura ambiente, mentre le temperature del suolo nudo supereranno l’ambiente fino a 15°C (Soil Conservation Service unpublished records, 1960s–1970s).,
Raccolta e germinazione dei semi
Semi maturi di 54 specie appartenenti a 16 famiglie e 37 generi sono stati raccolti tra gennaio e aprile 2009, 2010 e 2011 (vedi Tabella 1 per nomi completi e autorità. I buoni sono stati depositati presso l’Australian National Herbarium, Canberra). In totale la specie rappresentava più di un quarto della flora delle angiosperme australiane presenti nelle regioni alpine (Costin et al., 2000, vedi Tabella 1), anche se molte di queste specie si estendono anche al di sotto della linea degli alberi., La vitalità di tutte le collezioni è stata stimata prima della semina in condizioni di germinazione sperimentale utilizzando la tecnica di colorazione tetrazolium chloride (TZ) (International Seed Testing Association, 2003). Per maggiori dettagli sulla raccolta e l’elaborazione vedere l’Appendice A nel materiale supplementare.
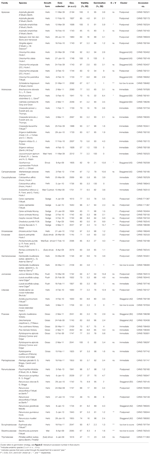
Tabella 1. Specie di studio, famiglia e autorità, forma di crescita e dettagli delle raccolte, percentuale di vitalità dei semi (TZ-stimata, ± s. e.), percentuale finale di germinazione raggiunta (±s. e.,), differenza statistica tra vitalità e germinazione (G < V, testato solo nei casi in cui la germinazione era inferiore alla vitalità).
La fenologia della germinazione dei semi è stata studiata imitando i regimi di temperatura che alpine seeds sperimenta in situ, post-dispersione, in una progressione artificialmente abbreviata delle stagioni. I semi sono stati assorbiti durante l’esperimento, quindi i risultati indicano il potenziale di germinazione quando l’acqua non è limitante., I dati sulla temperatura dell’aria e del suolo (Figura 1) sono stati utilizzati per guidare e convalidare i regimi di temperatura dell’incubatore, sebbene i vincoli logistici del lavoro con gli incubatori impedissero l’incorporazione delle fluttuazioni inerenti alle condizioni naturali. I test di germinazione hanno utilizzato otto repliche di 25 semi per raccolta, seminati in piastre di Petri di plastica del diametro di 9 cm contenenti l ‘ 1% di acqua naturale-agar. Le piastre di Petri sono state sigillate con parafilm per evitare l’essiccamento dell’agar, prima di essere collocate in incubatori di germinazione (Thermoline Scientific, Melbourne, NSW, Australia)., Metà dei piatti sono stati avvolti in un foglio di alluminio per escludere la luce e fornire un’indicazione dei requisiti di luce per la germinazione. Ciascuna delle quattro repliche per trattamento di luce per raccolta è stata collocata su un ripiano diverso all’interno della stessa incubatrice e la posizione all’interno di tale ripiano è stata riorganizzata settimanalmente. Tutte le repliche di ogni collezione sono state spostate attraverso i seguenti termoperiodi simulati: 8 settimane a 25/15°C (temperatura giorno/notte) → 2 settimane a 20/10 ° C → 2 settimane a 10/5 ° C → 10 settimane a costante 4-5 ° C → 2 settimane a 10/5 ° C → 2 settimane a 20/10 ° C → 6 settimane a 25/15°C., Pertanto, un ciclo dell’intero esperimento ha imitato le temperature che ricordano l’estate, l’inizio dell’autunno, il tardo autunno, l’inverno, l’inizio della primavera, la tarda primavera e l’estate, ed è durato 32 settimane in totale. L’anno abbreviato è stato considerato ragionevole perché le adesioni in genere avevano cessato un’ulteriore germinazione in un determinato trattamento prima di essere spostate alla “stagione”successiva., Un fotoperiodo chiaro/scuro 12/12 h è stato fornito da tubi fluorescenti (> 90 µmol m-2 s-1 al centro di ogni ripiano), e nei regimi di temperatura primaverile, estiva e autunnale la luce coincideva con il periodo di temperatura calda del giorno. La germinazione, definita come emergenza radicolare di oltre 1 mm, è stata valutata ogni 7 giorni e i semi e le piantine germinati sono stati successivamente rimossi dai piatti. La germinazione delle piastre avvolte in lamina è stata valutata come sopra, ma con luce verde molto bassa.,
Dopo un ciclo sperimentale (32 settimane) la maggior parte delle collezioni sono state terminate (vedi informazioni supplementari per le eccezioni), avendo mostrato zero germinazione per almeno 2 settimane in questo momento. Dopo la terminazione, tutti i semi rimasti intatti sono stati sezionati con un bisturi al microscopio. I semi con un endosperma e un embrione freschi e solidi sono stati considerati vitali e i semi vuoti di un embrione sono stati detratti dal totale quando si calcola la percentuale di germinazione per ogni raccolta., Abbiamo calcolato un indice dei requisiti di luce per la germinazione per ogni specie come frazione di germinazione nel trattamento scuro nel momento in cui sono stati raggiunti i percentili di germinazione cumulativi del 50 e del 95% nel trattamento alternato luce/buio (vedi sotto).
Correlati ecologici della strategia di germinazione
Abbiamo compilato un database di caratteristiche funzionali delle specie e misure geografiche dai dati pubblicati per valutare le correlazioni con la strategia di germinazione., Il database includeva i seguenti tratti continui: massa media del seme, requisito leggero per la germinazione (vedi sopra), area fogliare specifica (area fresca/massa secca), area fogliare individuale, peso fogliare individuale e altezza alla maturità. Le elevazioni minime, medie e massime di raccolta per la specie sono state determinate da tutti i record di erbario nell’erbario virtuale australiano, e l’elevazione a cui sono state fatte le raccolte di semi specifiche utilizzate nello studio è stata inclusa anche come tratti continui. La forma di crescita è stata valutata come una delle tre categorie: graminoide, erba o arbusto., Abbiamo anche incluso un tratto binario: se i semi fossero endospermici o meno.
I dati dei tratti sono stati raccolti da una varietà di fonti tra cui l’Australian Virtual Herbarium (http://avh.chah.org.au/), l’Australian National Herbarium specimen information register (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Alpine Flora (Costin et al., 2000), la flora del Nuovo Galles del Sud (Harden, 1990) e la morfologia interna comparativa dei semi (Martin, 1946). La massa del seme e l’area fogliare specifica sono state ottenute da singole raccolte di campo utilizzate nelle prove di germinazione., La massa del seme è stata ottenuta pesando quattro lotti di semi 10. SLA è stato determinato sulla base di cinque foglie completamente espanse da individui separati per specie. Questi sono stati raccolti sul campo, scansionati su uno scanner piano, essiccati a 60°C a peso costante e ponderati al più vicino 0.000 g. Dove erano disponibili semi insufficienti o non era disponibile un’area fogliare specifica, i dati sono stati ottenuti da altre raccolte ANBG della stessa specie o da altre fonti di dati (S. Venn, dati non pubblicati)., Se un seme fosse endospermico o meno è stato determinato dai disegni del seme nei registri della banca del seme di ANGB in consultazione con il personale della banca del seme di ANBG. Il database completo e le fonti sono disponibili su richiesta degli autori.
Analisi dei dati
I modelli di germinazione sono stati valutati visivamente e utilizzando un’analisi cluster in PC-ORD per Windows, versione 6 (MJM Software Design, Gleneden Beach, OR, USA). L’analisi ha utilizzato una matrice di distanza euclidea e un algoritmo di unione più vicino., I punti dati per l’analisi del cluster sono stati il tempo (settimane) a percentili di germinazione cumulativi specifici (t20, t40, t50, t60, t80 e t95%) per ogni raccolta (media tra i 4 piatti). Pertanto, una collezione che germinasse all’inizio dell’esperimento avrebbe tempi brevi per i percentili fino a t80%, ad esempio, mentre una collezione che ha posticipato la germinazione fino all’aumento delle temperature dopo l’inverno simulato avrebbe tempi lunghi per questi percentili., Modelli di germinazione più complessi, come la germinazione lenta e costante o esplosioni di germinazione, potrebbero anche essere rilevati e sarebbero indicati da tempi o tempi equidistanti raggruppati rispettivamente all’inizio e alla fine dell’esperimento. Questo metodo è stato utilizzato come un modo per imporre una valutazione oggettiva del modello di germinazione, indipendente dalla temperatura di per sé.,
È stata effettuata un’analisi unidirezionale della varianza (ANOVA) per le piastre replicate di ciascuna raccolta per valutare se vi fosse una differenza significativa tra la vitalità percentuale (determinata dal test TZ) e la germinazione percentuale finale. Ulteriori modelli lineari generali sono stati utilizzati per valutare se i cluster risultanti dall’analisi dei cluster differivano per vitalità o percentuale di germinazione finale.
Le associazioni tra tratti ecologici e cluster di strategia di germinazione sono state testate in due modi., In primo luogo, abbiamo testato le differenze medie tra cluster usando ANOVA con cluster come effetto fisso e correlati ecologici come variabili di risposta, o usando test G replicati per l’endospermia e la forma di crescita. In secondo luogo, poiché la germinazione e altri tratti funzionali sono altamente suscettibili di essere influenzati dalla storia evolutiva, abbiamo valutato queste correlazioni usando regressioni filogenetiche. Una filogenesi è stata costruita per le specie di studio utilizzando Phylocom Phylomatic e l’imballaggio bladj in R (Webb et al., 2008; R Core Team, 2013)., Abbiamo valutato la presenza della struttura filogenetica nelle variabili usando il phylosig o phylo.d (variabili discrete o binarie) funzione del pacchetto Fitools in R (Revell, 2012). Le regressioni filogenetiche sono state condotte utilizzando la funzione pgls in Caper (Orme et al., 2013). Le analisi sono state condotte in modo strutturato riflettendo il clustering rivelato in PC-Ord (vedi sotto)., Inizialmente abbiamo incluso quattro strategie di germinazione (categorie posticipate, immediate e scaglionate all’interno di ciascuna delle precedenti), poi tre (raggruppando tutte le specie scaglionate e confrontandole con posticipate e intermedie) e infine abbiamo valutato le differenze tra le due strategie più ampie, posticipate e intermedie.
Risultati
Vitalità dei semi Rispetto alla germinazione finale
Nel complesso, la vitalità dei semi e la percentuale di germinazione finale erano elevate. La redditività media stimata in TZ in tutte le collezioni è stata di 80 ± 1.,7%, con più di due terzi delle collezioni che mostrano una redditività superiore al 75% (Tabella 1). Non vi è stata alcuna differenza nella qualità delle raccolte nei 3 anni (la redditività media nel 2009, nel 2010 e nel 2011 era rispettivamente di 79 ± 2,8, 80 ± 2,7 e 81 ± 3,7%), indicando che il sistema bancario delle sementi raccolte nel 2009 non ha ridotto la loro redditività.
La germinazione che ha superato o non è stata significativamente inferiore alla vitalità stimata da TZ è stata raggiunta per 45 delle 54 specie (ANOVA a senso unico: P>0,05, Tabella 1)., Delle restanti nove specie, la germinazione finale di quattro specie era molto bassa (< 5%), nonostante una buona vitalità (ca. 55-94%), e quindi non siamo stati in grado di trarre conclusioni sulle strategie di germinazione di queste specie. La mancata germinazione potrebbe riflettere che i nostri trattamenti non alleviavano la dormienza o che i semi non erano completamente maturi alla raccolta. I test Cut alla fine dell’esperimento hanno indicato che, in tutti i casi, la maggior parte dei semi non germinati conteneva un embrione sano e assorbito., È interessante notare che 34 specie hanno raggiunto una germinazione superiore alla vitalità stimata da TZ (Tabella 1), suggerendo che il test TZ ha comunemente sottovalutato la vitalità dei semi.
Cluster di germinazione
L’analisi dei cluster ha rivelato due cluster principali (A e B), con sottostruttura in ciascuno (Figura 2). Per la maggior parte delle specie (n = 21 specie di 10 famiglie), la germinazione è stata posticipata fino a quando durante un periodo di temperature invernali (5°C giorno/notte) o fino a quando le temperature sono state elevate a 10/5°C dopo l’inverno (A1 e A1:inverno, Figure 2, 3A)., In alternativa, la germinazione è iniziata immediatamente a 25/15°C (n = 17 specie, sei famiglie, viste nel cluster B1, Figure 2, 3B). Per la maggior parte di queste specie, il tasso di germinazione è stato veloce; il 50% della germinazione totale (t50) si è verificato in meno di 3 settimane (Figura supplementare 1).
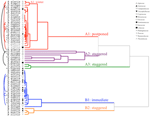
Figura 2. Analisi dei cluster dendrogramma basato sul tempo di percentili di germinazione cumulativa(20, 40, 50, 60, 80, e 95%). Specie indicata per genere iniziale e prima parte del nome della specie. Gli asterischi contrassegnano le raccolte che sono state bancate prima del test., Gli archi fini collegano le raccolte duplicate per confrontare l’effetto delle operazioni bancarie sulla germinazione. I simboli indicano famiglie diverse (vedi legenda). Le raccolte mostrate in testo grigio sono quelle che sono state esposte a due cicli dell’esperimento.
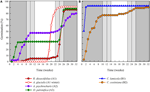
Figura 3. Percentuale cumulativa di germinazione di una specie rappresentativa di ciascun gruppo di germinazione. (A) cluster A, specie che ha posticipato la germinazione, (B) cluster B, specie che hanno germinato immediatamente., Si noti che i modelli di germinazione sfalsati sono nidificati all’interno dei cluster di cui sopra. I regimi di temperatura dell’incubatore sono rappresentati dall’ombreggiatura (vedi Figura 1).
All’interno di ciascuna di queste grandi classi, tuttavia, c’erano sotto-cluster chiari che riflettevano la germinazione che era sfalsata nel tempo, cioè una percentuale del lotto di semi germogliava prima dell’esposizione alle temperature invernali mentre il resto germogliava dopo l’inverno. La germinazione sfalsata si è verificata in 13 specie di sei famiglie e si è verificata principalmente quando le temperature diurne hanno superato i 10°C (Figure 3A,B)., Questi germinatori sfalsati sono stati suddivisi in tre gruppi nell’analisi del cluster. Il cluster A2 con sette collezioni ha mostrato una germinazione sostanziale sia prima che dopo il periodo di temperatura fredda. Dove la germinazione cessava prima che le temperature venissero cambiate, abbiamo concluso che la mancanza di un’ulteriore germinazione pre-invernale non era determinata dalla temperatura (A3). Al contrario, la germinazione di gran parte del gruppo A2 sembrava essere interrotta solo quando le temperature erano ridotte a 10/5°C., Il cluster B2 conteneva altre quattro specie che hanno mostrato una germinazione sostanziale all’inizio dell’esperimento e hanno continuato a raggiungere una percentuale relativamente piccola della loro germinazione totale dopo che le temperature sono state riportate sopra i 5°C (Figura 3B).
Rispetto alla variazione relativamente minore tra le specie all’interno dei cluster A1 e B1, i cluster sfalsati mostrano anche una differenziazione molto maggiore tra le specie. Le singole curve per tutte le specie sono riportate nelle figure supplementari 1i-vii. Per dettagli più specifici sulla strategia di germinazione vedere l’Appendice B nel Materiale supplementare.,
Ecologici Correlati di Germinazione Strategia
Di tutte le ecologica correla che abbiamo preso in considerazione, solo a coloro che sono direttamente associati con seme tratti erano significativamente correlati con la germinazione strategia che sia valutata la correlazione a livello di due cluster principali (rinviata vs immediata di germinazione), o a livello di tre o quattro cluster, o se non si hanno rappresentato per la struttura filogenetica o meno (Tabella 2)., Le specie con germinazione posticipata avevano semi più pesanti e avevano maggiori probabilità di essere endospermiche rispetto a quelle delle specie che germogliavano immediatamente (Figura 4). Le specie con germinazione sfalsata erano intermedie nella massa dei semi e nell’endospermia. Tuttavia, non tutte le caratteristiche dei semi erano correlate con la strategia di germinazione: non vi era alcuna correlazione tra i requisiti di luce per la germinazione dei semi e la strategia. Contrariamente alle nostre aspettative, le specie con un’area fogliare specifica più elevata (potenzialmente indicativa di tassi di crescita più elevati) non avevano maggiori probabilità di mostrare una germinazione immediata., Allo stesso modo, le specie con elevazione media più elevata o intervalli di elevazione più piccoli non avevano maggiori probabilità di mostrare una germinazione posticipata.

Tabella 2. Incidenza della struttura filogenetica in, e correlati di, strategia di germinazione.
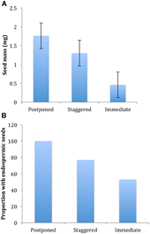
Figura 4., Differenze nella (A) massa media dei semi e (B) proporzione di specie con semi endospermici tra specie con strategie posticipate (cluster A1), sfalsate (combinazione di cluster A2, A3 e B2) e di germinazione immediata (cluster B1). Le differenze tra le strategie sono significative a P ≤ 0,05, vedi Tabella 2.
Discussione
La germinazione negli habitat alpini era storicamente considerata controllata dall’ambiente, con la neve invernale che isolava i semi dai potenziali segnali di germinazione e rendeva inutile la dormienza (Billings e Mooney, 1968)., Tuttavia, i nostri risultati supportano prove più recenti di meccanismi di dormienza comuni tra le specie alpine sia in Australia (Hoyle et al., 2013; Sommerville et al., 2013) e altrove in tutto il mondo (Cavieres e Arroyo, 2000; Schutz, 2002; Shimono e Kudo, 2005; Mondoni et al., 2009; Schwienbacher et al., 2011)., La dormienza sembrerebbe svolgere un ruolo significativo nel controllo dei tempi in situ della germinazione di molte specie alpine australiane, agendo per ritardare, posticipare o rallentare il tasso di germinazione e quindi potenzialmente conferire strategie di rigenerazione avverse al rischio in un ambiente duro e variabile. Forse la cosa più sorprendente dei nostri risultati di germinazione è stata la proporzione di specie per le quali la strategia di germinazione variava all’interno di una raccolta di semi per una data specie, suggerendo che entrambe le caratteristiche dormienti e non dormienti erano esposte all’interno della stessa raccolta di semi di queste specie., In particolare, le strategie di germinazione erano generalmente altamente conservate all’interno delle famiglie e anche correlate con elementi di anatomia del seme: massa ed endospermia. Le strategie di germinazione erano, tuttavia, indipendenti da altri tratti funzionali ecologicamente significativi della pianta matura.
Variazione nella strategia di germinazione
I semi di quasi la metà delle specie studiate sembravano incapaci di germinare fino a quando non erano esposti a un periodo fresco e umido (costante 5°C), suggerendo che la stratificazione a freddo alleviasse un meccanismo fisiologico di dormienza., Posticipare la germinazione fino alla primavera successiva può consentire alle piantine di evitare di stabilizzarsi sopra o prima del rigido inverno, ottimizzando al contempo la breve stagione di crescita imminente. Al contrario, la germinazione opportunistica esibita dalle specie che hanno germinato immediatamente può fornire un vantaggio selettivo quando il rischio di mortalità delle piantine invernali è basso, consentendo alle piante di fiorire prima della primavera successiva o di dimensioni maggiori (Donohue, 2002).,
Ad oggi, ci sono state poche prove pubblicate di variabilità intra-specifica nei meccanismi di dormienza all’interno delle collezioni di semi alpini, come spiegherebbe le strategie di germinazione scaglionate osservate qui. La germinazione sfalsata nel tempo può anche essere spiegata variando i livelli di maturità dei semi tra gli individui nella popolazione al momento della raccolta e/o potrebbe riflettere una strategia dimorfica all’interno di una singola pianta associata alla posizione sulla pianta o ai tempi di sviluppo.,
La copertura evolutiva delle scommesse è spesso evocata per spiegare la strategia diversificata nelle caratteristiche di germinazione dei semi (Simons, 2011), come quella dimostrata dalle collezioni con germinazione sfalsata. Se, tuttavia, una strategia diversificata si traduce sia in un aumento dell’idoneità media che in una riduzione della varianza (ad esempio, riducendo la concorrenza tra fratelli), allora non costituisce una copertura delle scommesse. Allo stesso modo, se l’apparente diversificazione riflette le differenze tra, non all’interno, gli individui, allora non è la copertura delle scommesse (Starrfelt e Kokko, 2012)., Le indagini sui modelli di dispersione delle piantine alpine e sulla tolleranza al gelo e allo scioglimento della neve, abbinate a studi sui determinanti della variazione della strategia di germinazione all’interno delle specie, possono aiutare a spiegare la fenologia della germinazione delle specie che presentano una strategia scaglionata e possono indicare se sono suscettibili di rappresentare strategie di copertura delle scommesse.,
Correlati ecologici della strategia di germinazione
Il nostro esame della relazione tra la strategia di germinazione e altri tratti ecologicamente importanti ha dimostrato che i tratti di germinazione erano correlati con altri tratti del seme ma non con i tratti vegetativi. Sommerville et al. (2013) ha scoperto che le erbe alpine australiane con semi non endospermici avevano maggiori probabilità di essere non dormienti alla dispersione. Il nostro lavoro supporta questa ricerca e indica inoltre che i semi più piccoli hanno maggiori probabilità di essere tra i germinatori immediati. Anche se Sommerville et al., (2013) non ha posto perché l’endospermia possa essere associata alla dormienza, suggeriamo che il posizionamento delle riserve nei cotiledoni possa migliorare il tasso di crescita precoce e l’insediamento in specie germinanti immediate. I nostri risultati non hanno indicato alcuna correlazione tra requisiti di luce e dormienza, sebbene i requisiti di luce variassero tra le specie. Ci sono una varietà di altri tratti di semi che tamponano il rischio di estinzione in ambienti variabili, ad esempio i meccanismi di dispersione dei semi e la longevità dei semi nella banca dei semi del suolo (vedi Tielborger et al., 2012 per una revisione)., In questa fase non possiamo dire se questi tratti mostrerebbero correlazioni con la strategia di germinazione o meno.
Abbiamo inoltre chiesto se la strategia di germinazione sarebbe correlata con tratti non riproduttivi ecologicamente importanti, come l’area fogliare specifica o l’altezza della pianta (Westoby, 1998; Wright et al., 2004). In particolare abbiamo ipotizzato che le specie germinanti immediate potrebbero mostrare valori di tratto indicativi di tassi di crescita più rapidi (utilizzando SLA come proxy) come potrebbe essere necessario per la creazione nel breve periodo tra la dispersione e l’inizio dell’inverno., Tuttavia, non abbiamo trovato tali correlazioni. Se i tratti fisiologici e di crescita delle piantine nelle prime fasi di stabilimento potrebbero mostrare modelli diversi rispetto ai tratti vegetativi adulti è una questione degna di considerazione, ma al di là dello scopo del presente lavoro.
Infine, abbiamo esaminato se le specie provenienti da altitudini più elevate in cui le stagioni di crescita sono generalmente più brevi potrebbero avere maggiori probabilità di esibire dormienza, ma non vi era alcuna associazione tra l’intervallo di elevazione, le elevazioni di raccolta o l’elevazione media della raccolta per la specie nel suo complesso e la strategia di germinazione., Insieme, questi risultati indicano che, mentre l’evoluzione della strategia di germinazione è strettamente legata a caratteristiche ecologicamente importanti del seme, comprende un asse in gran parte indipendente dalle caratteristiche vegetative o distributive.
Implicazioni per le comunità vegetali alpine in un clima che cambia
Gli ecosistemi alpini australiani, come le aree alpine di tutto il mondo, sono minacciati dai cambiamenti climatici combinati con i cambiamenti nella frequenza degli incendi, i modelli di utilizzo del suolo, l’afflusso di specie invasive e gli impatti di una maggiore visita umana., Il riscaldamento associato al cambiamento climatico si sta verificando più rapidamente al di sopra della linea degli alberi che a quote più basse, e si prevede che le aree alpine continueranno a sperimentare un riscaldamento superiore alla media in futuro (Kullman, 2004). È probabile che la capacità di continuare la rigenerazione attraverso le sementi in condizioni nuove svolga un ruolo significativo nella risposta delle piante alpine ai cambiamenti climatici e ai cambiamenti associati. Inoltre, è probabile che i regimi di disturbo alterati richiedano un ruolo maggiore di ripristino e riabilitazione nella regione., Le informazioni fornite qui sono quindi cruciali per la conservazione e la gestione dei sistemi alpini in fase di cambiamento; possono aiutare a informare le nostre previsioni su come le specie di studio possono essere influenzate, costituire la base dei piani di propagazione dei semi per queste specie e essere utilizzate per guidare le future indagini sulle strategie di germinazione dei semi alpini con,
I cambiamenti di temperatura possono influenzare lo stato di dormienza dei semi sia pre-che post-dispersione, alterando e/o interrompendo di conseguenza le strategie di germinazione e determinando cambiamenti nella fenologia di germinazione corrente (Hoyle et al., 2008b). Per le piante alpine, il riscaldamento climatico può portare a uno spostamento dalla primavera all’emergenza autunnale, guidato principalmente da cambiamenti nello stato di dormienza dei semi e con conseguenti importanti implicazioni per le specie attualmente adattate all’emergenza in primavera (Mondoni et al., 2012)., Ulteriori ricerche su come i cambiamenti nelle temperature che i semi sperimentano post-dispersione, la ridotta durata del manto nevoso e l’aumento della frequenza di eventi di temperatura estrema influenzeranno la germinazione sono necessarie per prevedere come le specie con dormienza risponderanno ai cambiamenti climatici. Per quelle specie alpine che germinano in autunno, prima del gelo e delle nevicate, i nostri risultati sollevano la questione di come le piantine di queste specie faranno fronte alla prevista riduzione delle precipitazioni e all’aumento della frequenza degli eventi di gelo stagionale.,
Infine, i nostri risultati sollevano importanti questioni riguardanti la variazione all’interno delle specie, in particolare in associazione con la previsione dell’impatto dei cambiamenti climatici o la selezione di quali semi utilizzare per il restauro. Le strategie di germinazione possono variare all’interno delle specie a seconda della popolazione, del tempo di fioritura, della massa dei semi, dell’investimento parentale, del clima, dell’elevazione e/o dell’ambiente materno e questa variazione può riflettere fattori genetici, epigenetici e/o ambientali (Cochrane et al., 2015)., L’elevazione crescente rappresenta la variazione dello stato di germinabilità e / o dormienza di diverse specie alpine (Billings e Mooney, 1968; Dorne, 1981; Cavieres e Arroyo, 2000) ma vedi (Hoyle et al., 2014). Lo stato di dormienza ridotto può derivare dalla cessazione anticipata dello sviluppo del seme derivante da stress ambientali (Steadman et al., 2006; Hoyle et al., 2008b)., L’esame delle variazioni nelle strategie di germinazione all’interno delle specie può rivelare il potenziale per le specie di far fronte e adattarsi ai cambiamenti climatici, svolgendo quindi un ruolo importante nel determinare la sopravvivenza futura e la distribuzione delle specie e fornendo informazioni importanti per la gestione e la conservazione.
Conclusioni
Il nostro approccio ha rivelato la presenza diffusa di dormienza nella flora alpina australiana e dimostra che le strategie di germinazione variano all’interno di una raccolta di semi, così come tra le collezioni., Inoltre, abbiamo dimostrato che mentre la strategia di dormienza è filogeneticamente conservata e correlata con la massa del seme e l’endospermia, è in gran parte indipendente dai tratti vegetativi e dalle caratteristiche della gamma. Mentre non si può estrapolare direttamente dai nostri risultati come il clima e i cambiamenti associati influenzeranno la flora alpina, questi risultati possono costituire la base per la progettazione di piani di propagazione e ulteriori sperimentazioni sull’impatto dei cambiamenti climatici., In particolare, sosteniamo ulteriori ricerche per comprendere le cause della variazione all’interno delle specie nella strategia di germinazione e il ruolo che questo può svolgere nella capacità delle specie di resistere ai cambiamenti climatici previsti.
Accessibilità dei dati
I dati completi di germinazione e il database dei tratti saranno depositati su DRYAD e con l’Atlante di Living Australia.
Dichiarazione sul conflitto di interessi
Gli autori dichiarano che la ricerca è stata condotta in assenza di relazioni commerciali o finanziarie che potrebbero essere interpretate come un potenziale conflitto di interessi.,
Riconoscimenti
Riconosciamo finanziamenti dall’Australian Research Council (LP0991593) e dall’Australian National Botanic Garden e dai suoi Amici. Ringraziamo J. McAuliffe, S. Fethers e S. Lawatsch dell’ANBG. Ringraziamo anche M. Holloway-Phillips, M. Cardillo, e S. Cunningham per l’input sulle versioni precedenti del manoscritto e per la consulenza analitica e S. Venn per l’accesso ai dati tratto.
Materiale supplementare
Foley, M. E. (1992). Effetto degli zuccheri solubili e dell’acido gibberellico nella rottura della dormienza degli embrioni di avena selvatica asportati (Avena fatua)., Weed Sci. 40, 208–214.
Google Scholar
Harden, GJ (1990). Flora del Nuovo Galles del Sud. Sydney, NSW: Royal Botanic Gardens.
Google Scholar
International Seed Testing Association. (2003). Fogli di lavoro sul test del tetrazolio. Bassersdorf: IST.
Kullman, L. (2004). Il volto mutevole del mondo alpino. Glob. Cambiare Newsl. 57, 12–14.
Google Scholar
Lascia un commento