
Il polimorfismo genetico è l’esistenza di stati alternativi del DNA, determinando la variazione dei livelli più alti di integrazione dell’organismo. Esistono diversi tipi di modificazioni del genoma (mutazioni). Le più studiate sono le sostituzioni nucleotidiche nelle regioni codificanti e regolatorie.
Definizione
Il polimorfismo genetico è l’esistenza, in una popolazione, di diversi stati alternativi di DNA, o alleli, in una posizione definita nel genoma, o locusPosition del gene sul cromosoma., In genetica di popolazione, un insieme di geni omologhi (classe di omologia). Due geni sono omologhi se corrispondono alla meiosi. Plurale: loci.. Questa definizione ha diversi aspetti:
(1) In primo luogo, il tratto deve essere trasportato dai cromosomi ed essere trasmissibile.
(2) Quindi gli alleli devono essere omologidue cromosomi o due geni si dicono omologhi se corrispondono e si escludono reciprocamente dalla meiosi. per la loro posizione nel genoma, che esclude alcuni tipi di variazione che saranno discussi in seguito.,
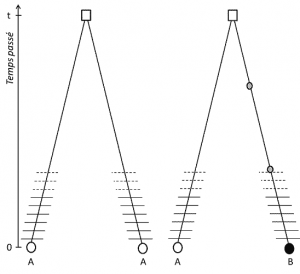
(3) Ma poiché il tratto è trasmissibile, l’omologia di posizione implica anche che gli alleli sono omologhi per discendenza; se sono diversi, è quindi che una mutazione (almeno) si è verificata in una delle linee che li collega al loro ultimo antenato comune (Figura 1).
(4) Infine, il polimorfismo genetico può essere definito alla scala della più piccola unità che compone il DNA: il sito nucleotidico., Di conseguenza, ogni variante nucleotidica può determinare il polimorfismo dei livelli più alti di struttura biologica-il gene, la proteina e il fenotipo dell’individuo-che diventano scale in cui il polimorfismo genetico può essere descritto (Figura 2).
Prendi l’esempio del sistema sanguigno ABO, che corrisponde a motivi antigeniciparte dell’antigene riconosciuto da un recettore anticorpale o linfocitario. Conosciuto anche come epitopo o determinante antigenico. Lo stesso antigene può avere diversi epitopi (identici o diversi) e quindi indurre una risposta immunitaria varia., coinvolto nella compatibilità durante le trasfusioni di sangue. Si tratta di un caso di polimorfismo genetico presente nelle popolazioni umane, che coinvolge la proteina ABO glicosil transferaseEnzima permettendo il trasferimento di residui contenenti zuccheri alle proteine. Nel caso del sistema ABO, la glicosiltransferasi A e B rispettivamente inducono un individuo ad appartenere al gruppo A o B. Quando entrambe le glicosiltransferasi sono presenti, l’individuo è del gruppo AB. che ha tre alleli, A, B e O. Un individuo può avere (AA), (AO); (BB), (BO); (OO) o (AB) genotipi., Il suo fenotipo sarà quindi,,;,; o, dove vediamo che A e B “dominano” O (cioè la loro espressione maschera quella di O), e che sono co-dominanti tra loro (l’eterozigote tra A e B ha un’espressione riconoscibile,).
Un polimorfismo può essere descritto per primo alla scala della sequenza di DNA del locus che codifica per una proteina. Alcuni polimorfismi sono “sinonimi”, cioè non modificano la sequenza aminoacidica della proteina; sono spesso i polimorfismi più numerosi. Gli altri modificano gli amminoacidi e sono chiamati polimorfismi “sostitutivi”., I polimorfismi sostitutivi della glicosil transferasi ABO sono di due tipi: polimorfismi che modificano l’aminoacido ma non l’unità antigenica (non interferiscono con il fenotipo ABO degli individui); e altri polimorfismi sostitutivi, che determinano il fenotipo ABO delle caratteristiche osservabili di un individuo..
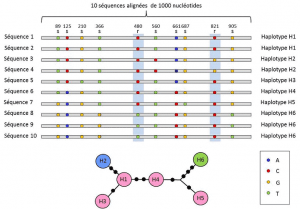
Tra i livelli di nucleotide e allele, le analisi genetiche tengono conto di un livello intermedio di descrizione: la disposizione lineare dei siti variabili sul locus. Queste disposizioni, chiamate aplotipigruppo di alleli di diversi loci situati sullo stesso cromosoma e di solito trasmessi insieme. L’aplotipo è un formato dalla contrazione della frase inglese genotipo aploide, o genotipo aploide., Tutti i geni situati sullo stesso cromosoma e i cui alleli si segregano insieme durante la meiosi costituiscono un aplotipo. Questi geni sono chiamati “geneticamente collegati”, sono utili nella genetica evolutiva perché permettono di trovare i collegamenti genealogici tra gli alleli (Figura 2).
I polimorfismi nucleotidici esistono anche nelle regioni intergeniche (al di fuori delle regioni codificanti). Alcuni influenzano le aree in cui l’espressione genica è regolata e quindi hanno espressione fenotipica. Altri non hanno alcun effetto noto e sono indicati come “silenziosi”.,
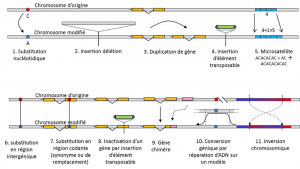
In aggiunta alle sostituzioni nucleotidiche, polimorfismo per un particolare tipo di mutazione, microsatelliti, si riferisce a variazioni nella durata di un’spesso non codificanti frammento di DNA (Figura 3) a causa di un cambiamento nel numero di ripetizioni in una breve sequenza ripetuta di nucleotidi, ad esempio, CACACACA, o TGTGTG.
Non tutte le variazioni genetiche rientrano nella definizione di polimorfismo genetico, poiché richiede l’omologia della posizione delle alternative (Figura 3)., Quindi, ci sono sequenze ripetute ottenute duplicando un gene, per il quale due ripetizioni non sono omologhe per posizione. Spesso, non possiamo parlare di omologia per gli elementi trasponibili, perché generalmente cambiano posizione durante le generazioni cellulari, possono moltiplicarsi e invadere il genoma. L’omologia di due elementi trasponibili non può quindi essere definita. D’altra parte, due sequenze codificanti dello stesso locus, una delle quali è intatta e l’altra inattivata dall’inserimento di un elemento trasponibile, corrispondono bene a una situazione di omologia., Un giorno, probabilmente considereremo di parlare di polimorfismo per le modifiche dei cosiddetti cromosomi “epigenetici”, che a volte vengono trasmessi su diverse generazioni cellulari, somaticiqualifica le cellule non riproduttive o soma. Una mutazione somatica, che colpisce un gene di una cellula somatica, scompare con l’individuo portatore. o germinalequalifica i gameti. Una mutazione germinale può essere trasmessa alla sua prole., come metilazione. La loro importanza nell’evoluzione delle popolazioni rimane da valutare., La limitazione della nozione di polimorfismo genetico a determinate categorie di variazione – essenzialmente sostituzioni nucleotidiche-deriva dalla grande utilità di quest’ultima nell’esplorare la storia delle popolazioni naturali. Si prestano alla modellazione matematica dell’evoluzione.
La parola polimorfismo significa “diverse forme”. Si oppone al monomorfismo, che indica l’assenza di variazione. Nel vocabolario della biologia, questa opposizione è stata a lungo utilizzata, prima della genetica, per segnalare la coesistenza di diversi tipi di individui nella stessa specie, ad esempio, polimorfismo di casta (regina e lavoratori) negli insetti sociali; polimorfismo stagionale (cambiamenti della pelliccia) in alcuni mammiferi artici. Questi casi non rientrano nel polimorfismo genetico, un concetto più recente con un significato più stretto e preciso. Anche le variazioni continue (ad esempio la variazione delle dimensioni) non rientrano nella categoria del polimorfismo, in quanto non presentano alternative distinte. Tuttavia, locus che influenzano la dimensione rientrano in questa definizione., Discuteremo di seguito le variazioni genetiche più studiate perché sono le più importanti per l’evoluzione, le sostituzioni nucleotidiche, e poi discuteremo la variazione dei fenotipi.
Misurazione
Non basta dire che un locus è più o meno variabile senza dare una stima quantitativa a questo giudizio. Un ricercatore può studiare la variazione a diverse scale. Se è interessato solo agli alleli di una proteina, misurerà la diversità allelica, simboleggiata da “H”. Se è interessato alla diversità del DNA, misurerà la diversità dei nucleotidi, simboleggiata da’π’.,
La diversità dell’allele H è definita come la probabilità di disegnare due alleli diversi durante due campionamenti con sostituzioneeseguire un sorteggio successivo con consegna di token p in un’urna contenente token n, significa prendere un primo token, leggere il suo valore, rimetterlo nell’urna, prendere un secondo token, leggere il suo valore, rimetterlo nell’urna, ecc., fino al token pth. Ciò significa scegliere gli oggetti p tra n con ripetizione (puoi scegliere lo stesso oggetto più volte) e in ordine (l’ordine in cui scegli gli oggetti è importante)., Il numero di estrazioni successive con token tra n è: n × n × n × × × n = np.. Se chiamiamo pi la frequenza dell’allele di rango i, mostriamo che la probabilità di raccogliere due volte lo stesso allele è F = Σ pi2. La diversità allelica del campione è quindi il suo complemento a 1, cioè:
H = 1 – Σ pi2 (1)
Questo vale sia per gli alleli proteici che per gli aplotipi. Questa è chiamata diversità aplotipica. Può anche essere chiamato eterozigosi, perché nel caso di un locus diploide, dà la frequenza prevista di eterozigoti.,
La diversità nucleotidica π è l’equivalente di H alla scala di ciascun nucleotide. Viene calcolato come la media del numero di differenze nucleotidiche tra sequenze in un campione prelevato due per due (δij), diviso per la lunghezza del frammento di DNA nel numero di nucleotidi (L).
π = media (δij) /L (2) (vedi riferimento)
Questo valore differisce tra le specie. Nelle parti codificanti del genoma umano, due cromosomi selezionati casualmente dalla popolazione differiscono in media di un nucleotide per mille., Nella mosca della frutta (Drosophila melanogaster) questa differenza è di circa uno su cento. La mosca è quindi dieci volte più variabile dell’uomo. Quando questi valori sono relativi alla dimensione delle regioni codificanti (circa 15.500 geni di Drosophila, almeno 22.000 in esseri umani), o anche genomi (140 milioni di coppie di basi per genoma aploide di mosche, circa 3.200 milioni di euro, venti volte di più nell’uomo), il numero di siti polimorfici è astronomico e risultati in nessun nati di sessualizzato generazione, in ogni specie è, è stato e continuerà ad essere geneticamente identico ad un altro passato, presente o futuro., Il potere informativo del polimorfismo del DNA è immenso. Nella scientifica, i detective possono identificare qualsiasi sospetto da sedici loculi di microsatellite.
Storia
Il termine variazione è nel titolo dei primi due capitoli del libro di Darwin The Origin of Species (1859). Darwin introdusse questa nozione nelle scienze naturali in un momento in cui le leggi dell’ereditarietà biologica rimasero un mistero e dedicò un altro importante libro ad esso, Variation in Animals and Plants (1868)., Convinto che l’evoluzione riguardasse piccole variazioni che facevano poca differenza per l’adattamento degli organismi alle loro condizioni di vita, attribuì importanza cruciale alle piccole variazioni quantitative, che spinsero i suoi successori (in particolare Karl Pearson) a fondare la biometria della misurazione della vita. Si riferisce in senso molto ampio allo studio quantitativo degli esseri viventi.. Ma la riscoperta delle leggi di Mendel nel 1900 spostò l’interesse verso variazioni discontinue.
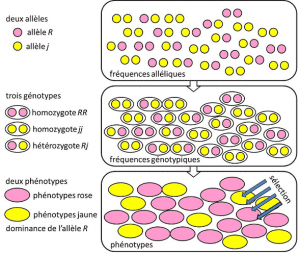
Dal 1908 al 1930, la genetica di popolazione era essenzialmente una disciplina teorica che cercava di conciliare il darwinismo con l’ereditarietà mendelianaeredità basata sulla trasmissione di un singolo gene sotto una modalità dominante, recessiva o legata al cromosoma sessuale X (o Y). Si riferisce a un tratto genetico con semplice determinismo, da una coppia o un piccolo numero di coppie di geni., e dove le probabilità hanno giocato un ruolo importante. La genetica è contro-intuitiva., Prevede che non vi sia riproduzione del genitore nel suo discendente, perché il genotipo di quest’ultimo risulta dalla lotteria della separazione degli alleli prima della fusione dei due semigenomi parentali. Si è capito che queste sono le frequenze allelichefrequenza alla quale l’allele di una variante si trova in una popolazione. Espresso in proporzione o percentuale. La somma delle frequenze alleliche di tutti gli alleli di un gene in una popolazione è quindi per definizione uguale a 1. Nella genetica delle popolazioni, le frequenze degli alleli rappresentano la diversità genetica a livello di popolazione o di specie., che vengono trasmessi tra generazioni, non genotipi o fenotipi. Queste frequenze sono più o meno stabili da una generazione all’altra e producono, al di là dell’intreccio multiplo di filiazioni tra individui, le stesse frequenze genotipichestruttura genetica della popolazione. Determinato dalle frequenze alleliche (Figura 4). La distribuzione dei genotipi di popolazione, associata alla varianza, è quindi l’unico fattore prevedibile., Intorno al 1930, i tre teorici Ronald Fisher, JBS Haldane e Sewall Wright contribuirono a creare la nozione della popolazione mendelianapopolazione la cui eredità obbediva alle leggi di Mendel. . In questo contesto, l’evoluzione associa necessariamente tre livelli strutturali: il gene, l’individuo e la popolazione. Questa associazione a tre vie può essere riassunta dalla seguente formula: l’evoluzione della popolazione è un cambiamento nella frequenza degli alleli (scala genica, ma anche scala di gruppo), un cambiamento che dipende dall’ordinamento dei fenotipi per selezione (scala individuale)., La ricerca empirica sulla genetica delle popolazioni si è poi sviluppata, ma è stata limitata a lungo dalla nostra ignoranza sul funzionamento dei cromosomi (la struttura del DNA è stata compresa nel 1953 e il suo sequenziamento è iniziato nel 1977), e ha dovuto ripiegare sullo studio dei pochi polimorfismi visibili, come la colorazione delle elitre di coccinella o il guscio delle lumache (vedi Focus The great snail debate). C’era molto dibattito tra i genetisti sul fatto che le popolazioni naturali fossero generalmente polimorfiche o monomorfiche e se il polimorfismo fosse di per sé benefico o meno., Lo studio della variazione della scala macromolecolare ha dovuto attendere fino al 1966 (per le proteine) e al 1983 (per il DNA) prima di rispondere a queste domande.
Variazioni quantitative e ereditabilità
Quando ci guardiamo intorno, vediamo differenze fisiche tra gli esseri, alcune complesse come i tratti del viso, altre facili da misurare come il peso o l’altezza. Il buon senso suggerisce che sono parzialmente trasmissibili, anche se in un modo difficile da specificare. Questi tratti sono spesso poligenicisotto la dipendenza di molti geni. Stiamo parlando di ereditarietà poligenica. Il diabete è una malattia poligenica.,, il che significa che sono influenzati da molti locus. La genetica dello sviluppo e la sua applicazione al confronto delle specie (evo-devo, per “evoluzione dello sviluppo”) districano la rete di interazioni complesse che fanno sì che durante lo sviluppo le cellule del corpo, tutte con identico bagaglio genetico, si differenziino per epigenesi per dare tessuti diversi. Non è ancora chiaro come siano costruiti personaggi complessi. I fenotipi misurabili possono essere studiati utilizzando la biometria, basata sulle proprietà delle distribuzioni statistiche., Un valore con proprietà utili è la varianzaIn un campione di individui in cui è stato misurato un dato tratto, la varianza è la differenza tra la media del quadrato dei valori e il quadrato della media dei valori. Questa misura, che è sempre positiva, indica la dispersione degli individui. . Quando diverse cause indipendenti determinano la variazione di una linea, le loro varianze sono additivi e la loro somma dà la varianza della linea. Se non sono indipendenti, la somma delle covarianze viene aggiunta alla somma delle varianze., La varianza totale Vt di una caratteristica fenotipica è tale che :
Vt = Vga+Vgd+Vgd+Vgi+Ve
Vga, genetico additivo varianza, è la somma delle varianze per ogni locus indipendentemente dagli altri; Vgd, genetica dominante varianza, è il risultato dell’interazione di due alleli di uno stesso locus; Vgi, interazione varianza tra locus, risultati dal fatto che il locus di uno stesso individuo di interagire; Ve, varianza dell’ambiente, qui si presume indipendente di varianza genetica.,
La varianza genetica della dominanza è comune a due figli degli stessi genitori, perché condividono gli effetti della dominanza quando in un determinato locus hanno ricevuto gli stessi alleli da entrambi i genitori, cosa che accade a loro in un locus su quattro. Non condividono questi effetti con i loro genitori, quindi sono più “simili” tra loro di quanto non siano ai loro genitori, nonostante condividano così tanti geni l’uno con l’altro. Certo, un genitore può, per caso, aver ricevuto gli stessi due alleli di uno dei suoi figli in un determinato luogo., Questa possibilità dipende dalla frequenza degli alleli nella popolazione. Vediamo che il grado di somiglianza tra un bambino e il suo genitore per un dato tratto è una proprietà pubblica.
Più in generale, tutte le componenti della varianza genetica (Vga+Vgd+Vgi) possono variare tra le popolazioni. Nelle linee riproduttive, un caso comune è l’inbreeding, che esaurisce la varianza genetica: di conseguenza, un tratto può avere la stessa media in due popolazioni, ma avere una variazione genetica diversa. La capacità di un allevatore di selezionare un tratto è quindi una caratteristica della linea, non del tratto.,
La relazione tra genitori e figli è misurata dall’ereditabilità h2. È definito come il rapporto tra varianza genetica additiva e varianza totale:
h2 = Vga/Vt

(2) Correlazione di 0, dove la dimensione dei bambini è indipendente da quella dei genitori. Un genitore medio di valore ha prodotto un figlio di valore indifferente, all’interno della distribuzione dei bambini. Le coordinate dei punti si trovano su un cerchio; l’aspettativa del loro valore si trova su una linea parallela all’asse dei genitori (d)., (3) Correlazione genitore-figlio intermedia tra 0 e 1: le coordinate dei punti si trovano su un’ellisse. L’aspettativa del loro valore non si trova più sulla bisettrice degli assi delle coordinate (che è anche l’asse maggiore dell’ellisse di distribuzione), ma sulla linea di regressione r. Un genitore medio di valore ha prodotto un figlio di valore b (tra 0 e a) con una certa varianza. Si noti che il caso (3) è intermedio tra i casi (1) e (2)
È mostrato sotto varie ipotesi nella Figura 5., Ereditabilità è la capacità di determinare se una caratteristica può essere selezionata. Figura 5-3 mostra che un allevatore sarà in grado, in un incrocio generazione, per spostare la media di un carattere dal valore 0 al valore di b se ha selezionato gli individui con un valore. Siamo in grado di mostrare che b = h2a. Per selezionare un tratto, è utile aumentare l’ereditabilità rapporto standardizzando l’ambiente (e quindi riducendo l’impatto ambientale della varianza), e da porsi in condizioni ambientali che favoriscono la comparsa della caratteristica., Il genetista Waddington ha dimostrato che, in natura, un cambiamento di ambiente può quindi rivelare nuovi tratti che verranno poi selezionati durante l’evoluzione.
Riferimenti e note
Immagine di copertina. Gusci di lumaca Cepaea nemoralis & Cepaea hortensis. André Künzelmann, UFZ.,
Questa formula può essere scritta in un modo più convenzionale come: \pi =\frac{n(n-1))}{2}\sum _{i=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
Essere in un campione di individui in cui l’studiato tratto stato misurato, la varianza può essere definito come la differenza tra la media dei quadrati dei valori e il quadrato della media dei valori. Questa misura, che è sempre positiva, indica la dispersione degli individui.
Questa è ereditabilità nel senso stretto del termine. L’ereditabilità in senso lato è il rapporto tra la somma delle varianze genetiche e la varianza totale., Questa formula dà ereditabilità con entrambi i genitori( il “genitore medio”); la stima con un solo genitore darebbe h2/2.
L’Enciclopedia ambientale dell’ambiente a cura dell’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contrattualmente legata all’Università di Grenoble Alpes e Grenoble INP, e patrocinata dall’Accademia francese delle Scienze.
Per citare questo articolo:VEUILLE Michel (2021), Genetic polymorphism and variation, Encyclopedia of the Environment, url:https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
Gli articoli dell’Enciclopedia dell’Ambiente sono resi disponibili secondo i termini della licenza Creative Commons BY-NC-SA, che autorizza la riproduzione a condizione di: citare la fonte, non farne uso commerciale, condividere condizioni iniziali identiche, riprodurre ad ogni riutilizzo o distribuzione la menzione di questa licenza Creative Commons BY-NC-SA.
Lascia un commento