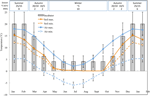
Figur 1. Månedlige maksimum og minimum luft-og jordtemperatur data i den Australske Alpene (gjennomsnitt ± STDEV). Lufttemperatur er gjennomsnitt fra BOM database, jord temperaturer ble målt med iButtons (se metoder). Linjene viser eksperimentelle inkubator temperaturer (gjennomsnitt ± STDEV)., Den tilsvarende tiden av året (sesong), inkubasjonstemperatur regimet (°C, 12/12 h, lys/mørk) og varighet av behandling (uker) som angitt i den øverste panel.
Med tanke på å avdekke temperaturforhold nær de som frø erfaring post-spredning, daglig maksimum og minimum jordtemperatur data ble samlet inn i studie-området (Sjømann Hytta; 36.27°S og 148.17°E, 2030 m en.s.l.) ved hjelp av ibutton dataloggere (n = 10, Innebygde datasystemer, USA) plasseres 4 cm under overflaten ved foten av vegetasjon fra 17. januar til 17 desember 2012., Vi loggere plassert på 4 cm for å unngå forstyrrelser på overflaten, men likevel være representative i forhold til frø som er eksponert i jord; denne dybden er middels til hva som har blitt brukt i tidligere alpine jord temperatur overvåking øvelser (Scherrer og Körner, 2011; Pauli et al., 2015). Som mistenkt, gjennomsnittlig jordtemperatur ikke falle under frysepunktet om vinteren (Figur 1). I sommer, jord temperatur under vegetasjonen er kjent for å spore omgivelsestemperatur, mens temperaturer på naken jord vil overstige ambient av opp til 15°C (Soil Conservation Service upubliserte registreringer, 1960–1970-tallet).,
Frø Innsamling og Spiring
Modne frø av 54 arter fra 16 familier og 37 arter ble samlet inn mellom januar og April 2009, 2010 og 2011 (se Tabell 1 for full navn og myndigheter. Bilag ble innlosjert på Australian National Herbarium, Canberra). I sum arter representert mer enn en fjerdedel av den Australske angiosperm flora funnet i alpine områder (Costin et al., 2000, se Tabell 1), selv om mange av disse artene omfatter nedenfor treeline som godt., Levedyktigheten til alle samlinger ble anslått før såing i eksperimentell spiring forhold ved å bruke den tetrazolium klorid (TZ) farging teknikk (International Seed Testing Association, 2003). For mer informasjon om innsamling og behandling se Vedlegg A i Supplerende Materiale.
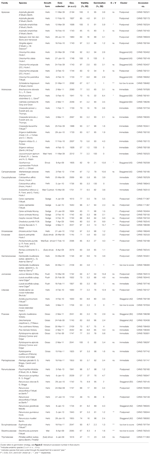
Tabell 1. Studien arter, familie og myndighet, vekst form, og samlinger detaljer, prosentandel spiredyktigheten (TZ-anslått, ± s.e.), final prosent spiring oppnådd (± s.e.,), statistisk forskjell mellom levedyktighet og tilvekst (G < V, testet kun i de tilfeller hvor spiring var mindre enn levedyktighet).
Phenology av frø spiring ble undersøkt ved å etterligne temperatur regimer som alpine frø erfaring in situ, post-spredning, i en kunstig forkortet progresjon av årstider. Frø ble imbibed hele eksperimentet dermed resultatene indikerer potensial for spiring når vann ikke er begrensende., Luft-og jordtemperatur data (Figur 1) ble brukt til å veilede og validere inkubator temperatur regimer, selv om den logistiske contraints av å arbeide med inkubatorer til hinder for inkorporering av valutasvingninger knyttet til naturlige forhold. Spiring tester brukt åtte replikater av 25 frø per samling, som ble sådd i 9 cm diameter plast Petri retter som inneholder 1% rent vann-agar. Petri retter var forseglet med Parafilm å unngå agar uttørking, før de blir plassert i spiring inkubatorer (Thermoline Vitenskapelige, Melbourne, NSW, Australia)., Halvparten av maten som ble pakket inn i aluminiumsfolie for å utelukke lys og gir en indikasjon på lys krav til spiring. Hver av de fire replikater per lysbehandling per samling var lagt på en annen hylle i samme inkubator og plassering innenfor det sokkel ble re-arrangert ukentlig. Alle replikater av hver samling ble flyttet gjennom følgende simulert thermoperiods: 8 uker på 25/15°C (dag/natt-temperatur) → 2 uker på 20/10°C → 2 uker på 10/5°C → 10 uker på konstant 4-5°C → 2 uker på 10/5°C → 2 uker på 20/10°C → 6 uker på 25/15°C., Dermed er en syklus av hele eksperimentet etterlignet temperaturer som minner om sommer, tidlig høst, sen høst, vinter, tidlig på våren, sent på våren og sommeren, og varte i 32 uker totalt. Forkortet år ble ansett som rimelig grunn accessions generelt hadde sluttet ytterligere tilvekst i en gitt behandling før de blir flyttet til neste «sesong»., En 12/12 h lys/mørk photoperiod ble gitt gjennom av lysstoffrør (> 90 µmol m−2 s−1 i midten av hver hylle), og i løpet av våren, sommeren og høsten temperatur regimer lys falt sammen med den varme temperaturen perioden av dagen. Spiring, definert som radicle fremveksten av mer enn 1 mm, ble scoret hver 7 dager, og spirer frø og frøplanter ble senere fjernet fra den retter. Spiring av folie-pakket platene ble vurdert som over, men under svært lave grønt lys.,
Etter en eksperimentell syklus (32 uker) de fleste samlingene var avsluttet (se supplerende informasjon for unntak), etter å ha vist null spiring i minst 2 uker på denne tiden. Følgende oppsigelse, alle gjenværende intakte frø ble dissekert med en skalpell under et mikroskop. Frøene med en fast, frisk endosperm og embryo ble ansett som forsvarlig og frø tomt av et embryo ble trukket fra det totale ved beregning av prosentandel spiring for hver samling., Vi beregnet en indeks av lys som trengs for spiring for hver art som er en brøkdel av spiring i mørket behandling på den tid som kumulative spiring prosentiler på 50 og 95% ble nådd i vekslende lys/mørk behandling (se nedenfor).
Økologiske Korrelerer av Spiring Strategi
Vi samlet en database of species » funksjonelle egenskaper og geografiske tiltak fra publiserte data for å vurdere sammenhenger med spiring strategi., Databasen inneholdt følgende kontinuerlig trekk: gjennomsnittlig frø masse, lys kravet for spiring (se ovenfor), bestemt blad området (frisk område/tørrvekt), individuelle blad området, individuelle blad tørr vekt og høyde ved forfall. Minimum, gjennomsnitt og maksimalt samling økninger for de artene som ble bestemt fra alle herbarium poster i den Australske Virtuelle Herbarium, og høyden på som den spesifikke frø samlinger brukt i studien ble gjort var også inkludert som kontinuerlig trekk. Veksten formen ble scoret som en av tre kategorier: graminoid, urt, eller busk., Vi har også inkludert en binær egenskap: om frø var endospermic eller ikke.
Den egenskap data ble samlet inn fra en rekke kilder, inkludert den Australske Virtuelle Herbarium (http://avh.chah.org.au/), Australian National Herbarium prøven informasjon register (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Alpin Flora (Costin et al., 2000), New South Wales Flora (Stivne, 1990), og Den komparative interne morfologi av frø (Martin, 1946). Frø masse og bestemte blad området ble innhentet fra de enkelte felt samlinger som brukes i spiring prøvelser., Frø masse ble innhentet ved å veie fire massevis av 10 frø. SLA ble fastsatt basert på fem fullt utvidet blader fra separate individer per art. Disse ble samlet inn i feltet, som er skannet på en flatbed scanner, tørket ved 60°C til konstant vekt og veid til nærmeste 0.000 g. Hvor utilstrekkelig frøet var tilgjengelig, eller spesifikke blad området ikke var tilgjengelig, data ble innhentet fra andre ANBG samlinger av samme art eller andre datakilder (S. Venn, upubliserte data)., Hvorvidt et frø var endospermic ble bestemt ut fra tegninger av frø i ANGB seed bank poster i samråd med ANBG frøbank ansatte. Full database og kilder som er tilgjengelige på forespørsel fra forfattere.
Data Analyse
Spiring mønstre ble vurdert visuelt og ved hjelp av en cluster-analyse i PC-ORD for Windows, versjon 6 (MJM Software Design, Gleneden Beach, ELLER, USA). Analysen brukes en Euclidean distance matrise og en nærmeste nabo bli med algoritmen., Data poeng for cluster-analysen var tid (uker) til bestemte kumulativ spiring prosentiler (t20, t40, t50, t60, t80, og t95%) for hver samling (gjennomsnitt over 4 retter). Dermed, en samling som spirer tidlig i eksperimentet ville har kort tid å prosentiler opp til t80% for eksempel, mens en samling som utsatt spiring til temperaturer økt etter simulert vinteren ville ha lange tider til disse prosentiler., Mer komplekse spiring mønstre, slik som langsom, jevn spiring eller utbrudd av spiring, kan også bli oppdaget, og vil bli indikert med jevnt fordelt ganger ganger eller gruppert i starten og slutten av forsøket, henholdsvis. Denne metoden ble brukt som en måte å innføre en objektiv vurdering av spiring mønster, uavhengig av temperatur per se.,
en En-veis analyse av varians (ANOVA) ble gjennomført for å kopiere plater av hver samling for å vurdere om det var en signifikant forskjell mellom andel levedyktighet (som bestemmes av TZ-test), og endelig prosent spiring. Ytterligere generelle lineære modeller ble brukt til å vurdere om klynger som følge av cluster-analyse ulik i levedyktighet eller siste tilvekst i prosent.
Assosiasjoner mellom økologiske egenskaper og spiring strategi klynger ble testet på to måter., For det første, vi testet for mener forskjellene mellom klynger ved hjelp av ANOVA med klyngen som en fast effekt og økologiske korrelerer som svar variabler, eller ved hjelp av replikert G-tester for endospermy og vekst form. For det andre, fordi spiring og andre funksjonelle egenskaper er svært sannsynlig å være påvirket av evolusjonære historie, har vi vurdert disse sammenhenger ved hjelp av fylogenetisk regresjoner. En fylogeni ble bygget for studier arter ved hjelp av Phylocom Phylomatic og bladj pakking i R (Webb et al., 2008; R Core Team, 2013)., Vi vurdert tilstedeværelsen av fylogenetisk struktur i variabler ved hjelp av phylosig eller phylo.d (diskret eller binære variabler) funksjon av Phytools pakke i R (Revell, 2012). Fylogenetisk regresjoner ble utført ved hjelp av pgls funksjon i Caper (Orme et al., 2013). Analysene ble gjennomført på en strukturert måte reflekterer clustering avslørt i PC-Ord (se nedenfor)., I utgangspunktet har vi inkludert fire spiring strategier (utsatt, umiddelbar, og sjanglet kategorier innenfor hvert av de foregående), deretter tre (lumping alle forskjøvet arter og sammenligne utsatt og intermediate) og til slutt har vi vurdert forskjeller mellom de to bredeste strategier, utsatt og middels.
Resultater
spiredyktigheten i Forhold til Siste Tilvekst
Samlet, spiredyktigheten og siste tilvekst andelen var høy. Mener TZ-estimert levetid på tvers av alle samlinger var 80 ± 1.,7%, med mer enn to tredjedeler av samlinger som viser mer enn 75% levedyktighet (Tabell 1). Det var ingen forskjell i kvaliteten på samlinger over 3 år (gjennomsnitt levedyktighet i 2009, 2010 og 2011 var 79 ± 2.8, 80 ± 2.7, og 81 ± 3,7 prosent, henholdsvis), noe som indikerer at bank av frø innsamlet i 2009 gjorde ikke minske deres levedyktighet.
Spiring at overskredet eller ikke var betydelig lavere enn TZ-estimert levedyktighet ble oppnådd for 45 av 54 arter (One-Way ANOVA: P>0.05, Tabell 1)., Av de resterende ni arter, siste tilvekst av fire arter var svært lav (< 5%), til tross for god levedyktighet (ca. 55-94%), og dermed var vi ikke i stand til å trekke noen konklusjoner om spiring strategier for disse artene. Unnlatelse av å spire kunne reflektere enten at våre behandlinger ikke lindre dvalen eller at frøene ikke var helt moden på samling. Kutt tester på slutten av eksperimentet viste at i alle tilfeller, de fleste av fn-spirer frø inneholdt en sunn, imbibed av embryo., Interessant, 34 arter oppnådd spiring som var større enn TZ-estimert levetid (Tabell 1), noe som tyder på at TZ test ofte undervurdert spiredyktigheten.
Spiring Klynger
Cluster analyse avslørte to store grupper (A og B), med sub-strukturen i hver (Figur 2). For de fleste artene (n = 21 arter fra 10 familier), spiring ble utsatt til i løpet av en periode på vintertemperatur (5°C dag/natt) eller til temperaturer som var oppdratt til å 10/5°C påfølgende vinter (A1 og A1:vinter, Tall 2, 3A)., Alternativt, spiring begynte umiddelbart på 25/15°C (n = 17 arter, seks familier, sett i klyngen B1, Tall 2, 3B). For de fleste av disse artene, spiring pris var rask; 50% av total tilvekst (t50) forekom i mindre enn 3 uker (Supplerende Figur 1).
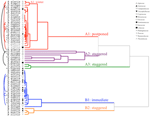
Figur 2. Cluster-analyse dendrogram basert på tide å kumulative spiring prosentiler (20, 40, 50, 60, 80, og 95%). Arter angitt med slekten første og første del av arter navn. Stjerner mark samlinger som ble samlet inn før testing., Fine buer koble dupliserte samlinger for å sammenligne effekten av banktjenester på spiring. Symboler betegne ulike familier (se tegnforklaring). Samlinger vist i grå tekst er de som ble utsatt for to sykluser av eksperimentet.
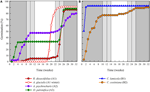
Figur 3. Kumulativ prosent spiring av én representant arter fra hver spiring klyngen. (En) klynge A, arter som utsettes spiring, (B) klynge B, arter som spirer umiddelbart., Vær oppmerksom på at forskjøvet spiring mønstre er nestet innenfor de ovennevnte grupper. Inkubator temperatur regimer er representert ved skyggelegging (se Figur 1).
Innenfor hver av disse brede klasser, men det var klart sub-klynger som reflekterer spiring som ble forskjøvet over tid, dvs. en andel av frø mye spirer før eksponering for vintertemperaturer mens resten spirer etter vinteren. Forskjøvet spiring skjedde i 13 arter fra seks familier og oppstod først og fremst når dagtemperaturer overskredet 10°C (figur 3A,B)., Disse forskjøvet germinators ble delt inn i tre grupper i cluster-analyse. Klyngen A2 med syv samlinger viste betydelig tilvekst både før og etter den kalde temperaturen periode. Hvor spiring sluttet før temperaturen blir endret vi konkluderte med at mangel på ytterligere pre-vinter spiring var ikke drevet av temperatur (A3). I kontrast, spiring av mye av A2-gruppen viste seg å bli stoppet bare når temperaturen ble redusert til 10/5°C., Klyngen B2 inneholdt en ytterligere fire arter som viste betydelig tilvekst tidlig i forsøket og gikk på å oppnå en relativt liten andel av deres totale tilvekst etter temperaturer ble returnert til over 5°C (Figur 3B).
i Forhold til det relativt liten variasjon mellom arter innenfor klynger A1 og B1, den forskjøvet klynger også viser mye større differensiering mellom arter. Enkelte kurver for alle arter er vist i Supplerende Tall 1i–vii. For mer spesifikke detaljer på spiring strategi se Vedlegg B i Supplerende Materiale.,
Økologiske Korrelerer av Spiring Strategi
Av alle økologiske korrelerer som vi vurderte, bare de som er direkte forbundet med frø trekk var signifikant korrelert med spiring strategi uavhengig av om vi vurdert korrelasjon på nivået av to store klynger (utsatt vs. umiddelbar spiring), eller på nivå med tre eller fire klynger, eller om vi sto for fylogenetisk struktur eller ikke (Tabell 2)., Arter med utsatt spiring hadde tyngre frø og var mer sannsynlig å være endospermic i forhold til de av arter som spirer umiddelbart (Figur 4). Arter med forskjøvet spiring var middels i frø masse og endospermy. Imidlertid, ikke alle frø egenskaper var korrelert med spiring strategi: det var ingen korrelasjon mellom lys som trengs for frø spiring og strategi. I motsetning til våre forventninger, arter med høyere spesifikke blad området (potensielt en indikasjon på høyere vekstrater) ikke var mer sannsynlig å vise umiddelbar spiring., Likeledes, arter med høyere gjennomsnittlig høyde eller mindre høyde varierer ikke var mer sannsynlig å vise utsatt spiring.

Tabell 2. Forekomsten av fylogenetisk struktur i, og korrelerer av, spiring strategi.
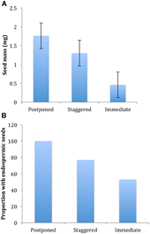
Figur 4., Forskjeller i (A) gjennomsnittlig frø masse og (B) andel av arter med endospermic frø blant arter med utsatt (cluster A1), forskjøvet (kombinasjon av klyngen A2, A3 og B2) og umiddelbar spiring (cluster B1) strategier. Forskjeller mellom strategier er signifikante ved P ≤ 0.05, se Tabell 2.
Diskusjon
Spiring i alpine habitater historisk var ansett for å være miljøvennlig kontrollert, med vinterens snø isolerende frø mot potensielle spiring signaler og gjengivelse av dvalen unødvendig (Billings og Mooney, 1968)., Men våre resultater gir støtte for flere nyere dokumentasjon av dvalen mekanismer for å være vanlig blant alpine arter både i Australia (Hoyle et al., 2013; Sommerville et al., 2013) og andre steder rundt om i verden (Cavieres og Arroyo, 2000; Schutz, 2002; Shimono og Kudo, 2005; Mondoni et al., 2009; Schwienbacher et al., 2011)., Dvalen synes å spille en betydelig rolle i å kontrollere in situ timing av spiring av mange Australske alpine arter, fungerende å utsette, utsette eller redusere frekvensen av spiring og dermed potensielt overdragelse risiko-uvillig regenerering strategier i en hard og varierende miljø. Kanskje mest slående av våre spiring resultatene var andelen arter som spirte strategi variert innenfor et frø samling for en gitt art som tyder på at både sovende og ikke-sovende egenskaper ble utstilt i samme frø samling av disse artene., Spesielt, spiring strategier var generelt svært bevart innenfor familier og også korrelert med elementer av frø anatomi: masse og endospermy. Spiring strategier var, imidlertid, uavhengig av andre økologisk betydelig funksjonelle egenskapene til den modne planten.
Variasjonen i Tilvekst Strategi
Frø av nesten halvparten av de artene som ble studert viste seg ute av stand til å spire før de ble utsatt for en kjølig og våt periode (konstant 5°C), noe som tyder på at kalde lagdelingen lindres en fysiologisk dvalen mekanisme., Å utsette spiring til våren kan aktivere seedlings å unngå å etablere over eller før den harde vinteren, mens også optimalisere kort kommende vekstsesong. I motsetning til den opportunistiske spiring utstilt ved arter som spirer umiddelbart kan gi en selektiv fordel når risikoen for vinteren frøplante dødeligheten er lav, ved at planter til å blomstre tidligere på våren eller ved en større størrelse (Donohue, 2002).,
hittil har det vært lite publiserte bevis av intra-spesifikke variasjon i dvalen mekanismer innen alpine frø samlinger som ville forklare forskjøvet spiring strategier observert her. Spiring forskjøvet over tid kan også forklares med varierende grad av frø modenhet blant personer i befolkningen på tidspunktet for samling, og/eller kunne reflektere en dimorphic strategi i et enkelt anlegg knyttet posisjon på anlegget eller timing av utviklingen.,
Evolusjonære bet-sikring er ofte fremkalt å forklare diversifisert strategi i frø spiring egenskaper (Simons, 2011), som demonstrert ved samlinger med forskjøvet spiring. Hvis, imidlertid, en diversifisert strategi resultater i både økte gjennomsnittlig fitness og en reduksjon i variansen (f.eks. ved å redusere konkurransen mellom søsken), så det utgjør ikke bet-sikring. Likeledes, hvis den tilsynelatende diversifisering gjenspeiler forskjeller blant, og ikke i, enkeltpersoner så det er ikke bet-sikring (Starrfelt og Kokko, 2012)., Undersøkelser i alpine frøplante spredning mønstre og toleranse for frost og snø smelter, sammen med studier av determinantene for spiring strategi variasjon innen artene, kan bidra til å forklare spiring phenology av artene viser en forskjøvet strategi og kan indikere om det er sannsynlig at de representerer bet-sikring strategier.,
Økologiske Korrelerer av Spiring Strategi
Vår undersøkelse av forholdet mellom tilvekst strategi og økologisk viktige trekk vist at spiring trekk var korrelert med andre frø trekk, men ikke med vegetative trekk. Sommerville et al. (2013) fant at Australske alpine urter med ikke-endospermic frø var mer sannsynlig å bli ikke-sovende på spredning. Vårt arbeid støtter at det å finne og videre indikerer at små frø er mer sannsynlig å være blant de umiddelbare germinators. Selv om Sommerville et al., (2013) ikke posit hvorfor endospermy kan være forbundet med dvalen, foreslår vi at plassering av reserver i cotyledons kan forbedre tidlig vekst og etablering i umiddelbar germinating arter. Våre resultater ikke viser noen sammenheng mellom lys krav og dvalen, selv om lyset gjorde krav varierer mellom arter. Det finnes en rekke andre frø egenskaper som buffer utryddelse risiko i variabel miljøer, for eksempel frø spredning mekanismer og frø levetid i jord frøbanken (se Tielborger et al. I 2012 for en gjennomgang)., På dette stadiet vi kan ikke si om disse trekkene ville vise korrelasjoner med spiring strategi eller ikke.
Vi har videre spurt om spiring strategi ville være korrelert med økologisk viktige ikke-reproduktive egenskaper, som for eksempel spesifikke blad området, eller plant høyde (Westoby, 1998; Wright et al., 2004). Vi spesielt hevdet at den umiddelbare germinating arter kan vise trekk verdier tyder på raskere vekst priser (ved hjelp SLA som en proxy) som kan være nødvendig for etablering i den korte perioden mellom spredning og utbruddet av vinteren., Men vi finner ingen slike sammenhenger. Om fysiologiske og vekst trekk av frøplanter i tidlig etablering stadier kan vise forskjellige mønstre enn voksne vegetative trekk er et spørsmål verdig betraktning, men utenfor rammen av det foreliggende arbeidet.
til Slutt, vi undersøkt om arter fra høyereliggende områder der vokser årstider er generelt kortere kan være mer sannsynlig å vise dvalen, men det var ingen sammenheng mellom enten høyde utvalg, innsamling høyder, eller gjennomsnittlig høyde på samling for arten som helhet og spiring strategi., Sammen er disse resultatene indikerer at mens utviklingen av spiring strategien er nært knyttet til økologisk viktige frø trekk, det består av en stor grad uavhengig aksen til vegetative eller distributional egenskaper.
Implikasjoner for Alpine Plante Samfunn i et Endret Klima
Australske alpine økosystemer, som alpine områder rundt om i verden, er under trusselen fra klimaendringer kombinert med endringer i brann-frekvens, arealbruk mønstre, tilstrømningen av fremmede arter, og virkningene av økt menneskelig samvær., Oppvarming forbundet med klimaendringene skjer raskere over treeline enn ved lavere høyder, og alpine områder er spådd til å fortsette å oppleve over gjennomsnittlig oppvarming i fremtiden (Kullman, 2004). Kapasiteten for fortsatt regenerering via frø under romanen forhold er sannsynlig å spille en betydelig rolle i svar av alpine planter til klima og tilhørende endringer. I tillegg, endret forstyrrelse regimer er sannsynligvis kreve en økt rolle for restaurering og rehabilitering i regionen., Den innsikt som er gitt her er derfor avgjørende for å bevare og forvalte alpine systemer under endring; de kan hjelpe med å informere våre spådommer om hvordan studere arter kan bli berørt, danner grunnlag av frø forplantning planer for disse artene, og brukes til å lede fremtidige undersøkelser i alpine frø spiring strategier med identifiserbare bevaring og forvaltning utfall.,
Endringer i temperatur er egnet til å påvirke dvalen status av frø både pre – og post-spredning, følgelig å endre og/eller avbryte spiring strategier og resulterer i endringer i gjeldende spiring phenology (Hoyle et al., 2008b). For alpine planter, klima oppvarming kan føre til et skifte fra høsten til våren dukker opp, drevet hovedsakelig av endringer i frø dvalen status og resulterer i store implikasjoner for arter tiden tilpasset fremveksten våren (Mondoni et al., 2012)., For mer forskning på hvordan endringer i temperaturer som frø erfaring post-spredning, redusert varighet av snødekke, og økt frekvens av ekstreme temperatur hendelser vil påvirke tilvekst er nødvendig for å kunne forutsi hvordan arter med dvalen vil reagere på klimaendringer. For de alpine arter som gror i høst, før frost og snøfall, våre resultater heve spørsmålet om hvordan frøplanter av disse artene vil takle den forventede reduksjon i nedbør og økt frekvens av a-sesongmessig frost hendelser.,
til Slutt, våre resultater opp viktige spørsmål om variasjon innen artene, spesielt i forbindelse med å forutsi effekter av klimaendringer eller velge hvilke frø til å utnytte for restaurering. Spiring strategier kan variere mellom arter avhengig befolkningen, blomstrende tid, frø masse, foreldrenes investeringer, klima, høyde, og/eller mors miljø, og denne varianten kan gjenspeile genetisk, epigenetisk, og/eller miljømessige faktorer (Cochrane et al., 2015)., Økende høyde kontoer for variasjon i germinability og/eller dvalen status for flere alpine arter (Billings og Mooney, 1968; Dorne, 1981; Cavieres og Arroyo, 2000), men se (Hoyle et al., 2014). Redusert dvalen status kan resultere fra tidlig avslutning av frø utvikling som følge av miljømessige stressfaktorer (Steadman et al., 2006; Hoyle et al., 2008b)., Undersøkelse av variasjonen i tilvekst strategier innen artene kan avdekke potensial for arter å takle, og tilpasse seg endringer i klimaet, og dermed, som spiller en viktig rolle i å bestemme fremtidig overlevelse og arter-distribusjoner, og gir viktig innsikt for forvaltning og bevaring.
Konklusjon
Vår tilnærming har avdekket utbredt forekomst av dvalen i den Australske alpin flora og viser at spiring strategier variere innenfor et frø samling, samt mellom samlingene., Videre har vi vist at mens dvalen strategi er phylogenetically bevart og korrelert med masse frø og endospermy, det er i stor grad uavhengig av vegetative egenskaper og utvalg egenskaper. Mens man kan ikke ekstrapolere direkte fra våre resultater hvordan klima og tilhørende endringer vil påvirke alpin flora, er disse resultatene kan danne grunnlag for utforming av forplantning planer og videre eksperimentering på virkningen av endrede klimatiske regimer., Spesielt kan vi argumentere for mer forskning på årsakene til innen-arter variasjon i tilvekst strategi og rolle dette kan spille på muligheten av arter til å tåle spådd klimaendringer.
Data Tilgjengelighet
Full spiring data og trekk database vil bli innlosjert på DRYAD og med Atlas of Living Australia.
interessekonflikt Uttalelse
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller finansielle forhold som kan oppfattes som en potensiell interessekonflikt.,
Erkjennelsene
Vi erkjenner finansiering fra den Australske forskningsrådet (LP0991593) og Australian National Botanic Garden og Venner av disse. Vi takker J. McAuliffe, S. Fethers, og S. Lawatsch av ANBG. Vi vil også takke M. Holloway-Phillips, M. Cardillo, og S. Cunningham for-inngang på tidligere versjoner av manuskriptet og for analytisk råd og S. Venn for tilgang til trekk data.
Supplerende Materiale
Foley, M. E. (1992). Effekten av løselig sukker og gibberellic-syre i å bryte dvalen av excised Vill Havre (Avena fatua) embryo., Luke Sci. 40, 208-214.
Google Scholar
Stivne, G. J. (1990). Flora of New South Wales. Sydney, NSW: Royal Botanic Gardens.
Google Scholar
Internasjonal Frø Testing Association. (2003). Arbeider Ark på Tetrazolium Testing. Bassersdorf: istanbuls iko.
Kullman, L. (2004). Endre ansiktet av alpine verden. Glob. Endre Newsl. 57, 12-14.
Google Scholar
Legg igjen en kommentar