
Genetiske polymorphism er eksistensen av alternative tilstander av DNA, for å fastslå variant av organismens høyere nivåer av integrasjon. Det finnes forskjellige typer av genom endringer (mutasjoner). Den mest studert er nukleotid innbytter i koding og regulatoriske regioner.
Definisjon
Genetiske polymorphism er eksistens, i en befolkning, av flere alternative tilstander av DNA, eller alleler, på en definert posisjon i genom, eller locusPosition av genet på kromosom., I populasjonsgenetikk, et sett av homologe gener (homologi klasse). To homologe gener er hvis de samsvarer meiose. Flertall: loci.. Denne definisjonen har flere aspekter:
(1) Først, trekk må utføres av kromosomene og smitte.
(2) Deretter alleler må være homologousTwo kromosomer eller to gener som er sagt å være homologe om de passer, og gjensidig utelukke hverandre fra meiose. for deres posisjon i genom, noe som utelukker visse typer av variasjon som vil bli diskutert senere.,
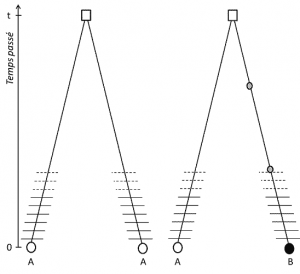
(3), Men siden den egenskap er overførbar, posisjon homologi innebærer også at alleler er homologe av nedstigningen, hvis de er forskjellige, det er derfor at en mutasjon (minst) har oppstått i en av de linjene som knytter dem til deres siste felles stamfar (Figur 1).
(4) til Slutt, genetisk polymorphism kan defineres på omfanget av den minste enheten du skriver DNA: den nukleotid nettstedet., Som et resultat, hver nukleotid variant kan bestemme polymorphism av høyere nivåer av biologisk struktur – den enkeltes genet, protein og fenotype – som blir skalaer som genetisk polymorphism kan være beskrevet (Figur 2).
Ta eksempel av ABO blod system, som tilsvarer antigene motivesPart av antigen anerkjent av et antistoff eller lymfocytt-reseptor. Også kjent som en epitope eller antigene determinanten. Det samme antigenet kan ha flere epitopes (identiske eller forskjellige) og dermed indusere en variert immunrespons., involvert i kompatibiliteten under blodoverføringer. Dette er et tilfelle av genetisk polymorphism til stede i menneskelige populasjoner, som involverer proteinet ABO glycosyl transferaseEnzyme å tillate overføring av reststoffer som inneholder sukker til proteiner. I tilfelle av ABO-systemet, glycosyl transferase A og B henholdsvis indusere en person hører til i gruppe A eller B. Når både glycosyltransferases er til stede, individet er av group AB. som har tre alleler, A, B og O. En person kan ha (AA), (AO); (BB), (BO); (OO) eller (AB) genotypes., Dens fenotype vil da bli,,; ,;, eller, der ser vi at A og B «dominere» O (dvs. deres uttrykk masker som O), og at de er co-dominant seg i mellom (den heterozygote mellom A og B har et gjenkjennelig uttrykk ).
En polymorphism kan beskrives først på omfanget av DNA sekvens av locus-koding for et protein. Noen polymorfismer er «synonymt med», dvs. at de ikke endrer aminosyresekvens av protein; de er ofte de mest tallrike polymorfismer. Den andre endre aminosyrer og er kalt «erstatning» polymorfismer., Erstatning polymorfismer av ABO glycosyl transferase er av to typer: polymorfismer som endrer aminosyre, men ikke antigene enhet (de ikke kommer i konflikt med ABO fenotypen av enkeltpersoner), og andre utskifting polymorfismer, som avgjør ABO phenotypeSet av observerbare kjennetegn ved en person..
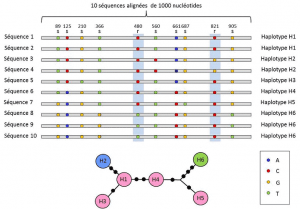
Mellom nukleotid og allelet nivåer, genetiske analyser som tar hensyn til et middels nivå av beskrivelse: lineær ordning med variabel nettsteder på locus. Disse ordningene, kalt haplotypesGroup av alleler av ulike loci som ligger på samme kromosom, og vanligvis overføres sammen. Haplotype er en dannet ved sammentrekning av det engelske uttrykket haploid genotype, eller haploid genotype., Alle gener som ligger på samme kromosom, og der allelene skiller sammen under meiose utgjør en haplotype. Disse genene kalles «genetisk koblet», er nyttig i evolusjonær genetikk, fordi de tillater å finne den genealogiske koblinger mellom alleler (Figur 2).
Nukleotid polymorfismer også finnes i intergenic regioner (utenfor koding regioner). Noen innvirkning på de områdene der genuttrykk er regulert, og har derfor fenotypiske uttrykk. Andre har ingen kjent effekt og er referert til som «stille».,
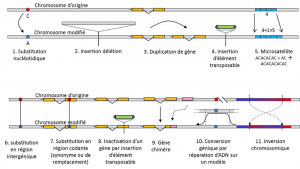
I tillegg til nukleotid erstatninger, polymorphism for en bestemt type mutasjon, microsatellites, viser til variasjoner i lengden på en ofte ikke-kodende DNA-fragment (Figur 3) på grunn av en endring i antall repetisjoner i en kort gjentatt sekvens av nukleotider, f.eks. CACACACA, eller TGTGTG.
Ikke alle genetiske variasjonen faller innenfor definisjonen av genetiske polymorphism, siden det krever stilling homologi av alternativene (Figur 3)., Dermed er det repeterte sekvenser som oppnås ved å duplisere et gen, som to repetisjoner er ikke homologe per stilling. Ofte, kan vi ikke snakke om homologi for transposable elementer heller, fordi de vanligvis endre posisjon under celle generasjoner, kan multiplisere og invadere genom. Den homologi av to transposable elementer ikke da være definert. På den annen side, to kodende sekvenser av samme locus, som er intakt, og den andre inaktivert ved innsetting av en transposable element, svarer godt til en homologi situasjon., En dag vil vi nok vurdere å snakke om polymorphism for endringer av såkalte «epigenetisk» kromosomer, som er noen ganger overført over flere celle generasjoner, somaticQualifies ikke-reproduktive celler, eller soma. En somatisk mutasjon, noe som påvirker et gen for en somatisk celle, forsvinner med transportøren enkelte. eller germinalQualifies kjønnsceller. En germline mutasjon kan overføres til sine avkom. slik som metylering. Deres betydning i utviklingen av bestander som gjenstår å bli vurdert., Begrensning av begrepet genetisk polymorphism til visse kategorier av variasjon – i hovedsak nukleotid innbytter – kommer fra den store nytten av det siste i å utforske historien til naturlige bestander. De egner seg til matematisk modellering av utviklingen.
ordet polymorphism betyr «flere former». Det er i motsetning til monomorphism, noe som indikerer fravær av variasjon. I vokabularet av biologi, denne opposisjonen har lenge vært brukt, før genetikk, for å signalisere sameksistens mellom flere typer av individer i de samme artene, f.eks., kastesystemet polymorphism (dronning og arbeidere) i sosiale insekter; sesongens polymorphism (pels endringer) i noen Arktiske pattedyr. Disse tilfellene faller ikke inn under genetisk polymorphism, et nyere konsept med et smalere og mer presis betydning. Kontinuerlig varianter (f.eks. størrelse variasjon) også ikke faller inn i den kategorien polymorphism, siden de ikke er til stede distinkte alternativer. Imidlertid, locus som påvirker størrelsen faller innenfor denne definisjonen., Vi vil diskutere nedenfor den mest studerte genetiske variasjoner fordi de er mest viktig for evolusjon, nukleotid-erstatninger, og så vil vi diskutere variant av phenotypes.
Måling
Det er ikke nok å si at et locus er mer eller mindre variabel uten å gi et kvantitativt estimat for denne dom. En forsker kan studere variasjon på ulike skalaer. Hvis han er bare interessert i alleler av et protein, han vil måle genetisk mangfold, symbolisert ved «H». Hvis han er interessert i DNA mangfold, han vil måle nukleotid mangfold, symbolisert ved ‘π’.,
allelet mangfold H er definert som sannsynligheten for å trekke to ulike alleler i løpet av to prøvetaking med replacementPerforming en påfølgende tegne med levering av s innsats i en urne inneholder n merker, betyr å ta et første tokenet, lesing sin verdi, sette det tilbake i urnen, tar en annen måte å lese sin verdi, sette det tilbake i urnen, etc. inntil pth-token. Dette betyr å velge p-objekter i n med repetisjon (du kan velge det samme objektet flere ganger) og i rekkefølge (den rekkefølgen du velger objektene er viktig)., Antall påfølgende trekker med tokens blant n) er: n × n × n × … × n = np.. Hvis vi kaller pi frekvensen av allelet av rang jeg, vi viser at sannsynligheten for å plukke to ganger samme allel er F = Σ pi2. Den genetisk mangfold for eksempel er da dens komplement til 1, dvs.:
H = 1 – Σ pi2 (1)
Dette gjelder både protein-alleler og haplotypes. Dette kalles haplotypical mangfold. Det kan også bli kalt heterozygosity, fordi i tilfelle av en diploid locus, det gir den forventede frekvensen av heterozygote katter.,
nukleotid mangfold π er tilsvarende H på omfanget av hvert nukleotid. Det er beregnet som gjennomsnittet av antall nukleotid forskjeller mellom sekvenser i en prøve tatt to og to (δij), dividert med lengden av DNA-fragment i antall nukleotider (L).
π = gjennomsnitt (δij)/L (2) (se referanse )
Denne verdien varierer mellom arter. I koding deler av det menneskelige genom, to kromosomer tilfeldig valgt fra befolkningen varierer i gjennomsnitt med ett nukleotid per tusen., I bananflue (Drosophila melanogaster) denne forskjellen er om en i hundre. Fly er dermed ti ganger mer variabel enn mannen. Når disse verdiene er knyttet til størrelsen av kodende regioner (om 15,500 gener i Drosophila, minst 22 000 i mennesker), eller til og med genomer (140 millioner base par per haploid genom i fluer, om 3,200 millioner kroner, eller tjue ganger mer på mennesker), antall polymorf nettsteder er astronomiske, og resulterer i at ingen blir født av seksualisert generasjon i noen arter er, har vært og vil være genetisk identisk med en annen en fortid, nåtid eller fremtid., Informasjonen kraft av DNA polymorphism er enorm. I kveld, detektiver kan identifisere noen mistanke fra seksten microsatellite locuses.
Logg på
begrepet variasjon er i tittelen på de to første kapitlene av Darwins bok The Origin of Species (1859). Darwin introduserte dette begrepet i realfag i en tid da lover biologiske arv forble et mysterium, og viet en annen viktig bok til det, Variasjon i Dyr og Planter (1868)., Overbevist om at utviklingen var i ferd med små variasjoner som gjorde liten forskjell til tilpasning av organismer til sine levekår, han festet avgjørende betydning for små kvantitative varianter, som førte til hans etterfølgere (særlig Karl Pearson) til fant biometryScience av livet måling. Refererer i en svært bred forstand til kvantitativ studie av levende vesener.. Men gjenoppdagelsen av Mendel lover i 1900 flyttet interesse for å usammenhengende variasjoner.
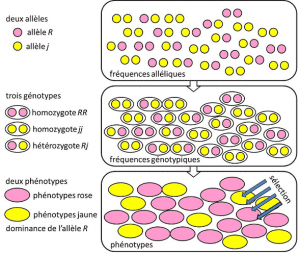
Fra 1908 til 1930, populasjonsgenetikk var i hovedsak en teoretisk disiplin søker å forene Darwinismen med Mendelian heredityHeredity basert på overføring av et enkelt gen under en dominerende, recessiv eller sex-kromosom-knyttet-modus X (eller Y). Refererer til en genetisk egenskap med enkle determinisme, ved et par eller et lite antall par av gener. og der sannsynlighetene spilte en viktig rolle. Genetikk er intuitiv., Det spår at det er ingen reproduksjon av morselskapet i hans etterkommer, fordi sistnevnte er genotype resultater fra lotteri av separasjon av alleler før fusjon av de to foreldrenes halv-genomet. Det ble innsett at dette er allel frequenciesFrequency som allelet av en variant er funnet i en befolkning. Uttrykt som andel eller prosent. Summen av allelet frekvenser av alle alleler av et gen i en populasjon er derfor per definisjon er lik 1. I populasjonsgenetikk, allelet frekvenser representerer det genetiske mangfoldet i befolkningen eller arter nivå., som overføres mellom generasjoner, ikke genotypes eller phenotypes. Disse frekvensene er mer eller mindre stabil fra en generasjon til den neste, og produsere, utover flere forviklinger av filiations mellom individer, den samme genotypic frequenciesGenetic struktur av befolkningen. Fastsatt av allel-frekvenser (Figur 4). Fordelingen av befolkningen genotypes, forbundet med avvik, er derfor den eneste forutsigbare faktor., Rundt 1930, tre teoretikere Ronald Fisher, JBS Haldane og Sewall Wright bidratt til å skape forestillingen om Mendelian populationPopulation som arvelighet adlød Mendel ‘ s lover. . I denne sammenheng er utviklingen nødvendigvis knytter tre strukturelle nivåer: genet, den enkelte og befolkningen. Dette tre-veis foreningen kan oppsummeres med følgende formel: befolkning evolusjon er en endring i allelet frekvens (gen skala, men også gruppe-skala), en endring som er avhengig av sortering av phenotypes av utvalget (individuelle skala)., Empiriske populasjonsgenetikk forskning deretter utviklet, men var begrenset i lang tid av vår uvitenhet om hvordan kromosomene arbeid (strukturen av DNA ble forstått i 1953 og dens sekvensering begynte i 1977), og måtte falle tilbake på studiet av de få synlige polymorfismer, for eksempel fargen på ladybug ‘ s elytra eller skallet av snegler (se Fokusere stor snegl debatt). Det var mye debatt blant genetikere som om naturlige bestander var generelt polymorfe, eller bare, og om polymorphism var i seg selv er gunstig eller ikke., Studiet av macromolecular skala variasjon måtte vente helt til 1966 (for proteiner) og 1983 (til DNA) før du svarer på disse spørsmålene.
Kvantitative variasjoner og arvbarhet
Når vi ser rundt oss, ser vi fysiske forskjeller mellom mennesker, noen komplekse som ansiktstrekk, andre lett å måle som vekt eller høyde. Sunn fornuft tilsier at de er delvis smitte, selv om det på en måte som er vanskelig å angi. Disse trekkene er ofte polygenicunder avhengighet av mange gener. Vi snakker om polygenic arvelighet. Diabetes er en polygenic sykdom., som betyr at de er påvirket av mange locuses. Utviklingsmessige genetikk, og sin søknad til sammenligning av arter (evo-devo, for «evolution of development») løse nettet av komplekse interaksjoner som gjør at det under utvikling celler i kroppen, alle med samme genetiske bagasje, differensiere seg ved å epigenesis å gi forskjellige vev. Det er fortsatt uklart hvordan komplekse karakterer er bygget. Målbare phenotypes kan bli studert ved hjelp av biometri, basert på egenskaper av statistiske fordelinger., Én verdi med nyttige egenskaper er varianceIn et utvalg av personer som i en gitt egenskap ble målt, variansen er forskjellen mellom gjennomsnittet av kvadratet av verdier og torget gjennomsnittet av verdiene. Dette tiltaket, som alltid er positiv, indikerer spredningen av enkeltpersoner. . Når flere uavhengige årsaker bestemme variant av en linje, deres avvik er additiv og deres sum gir variansen av linjen. Hvis de ikke er uavhengige, summen av covariances er lagt til summen av variansene., Den totale variansen Vt av en fenotypiske trekk er slik at :
Vt = Vga+Vgd+Vgd+Vgi+Ve
Vga, additiv genetisk variasjon er summen av variansene på grunn av hvert locus uavhengig av de andre, Vgd, dominerende genetisk varians, resultater fra samspillet av to alleler med samme locus; Vgi, samhandling avvik mellom locuses, resultater fra det faktum at locuses av samme individ samhandle; Ve, varians av miljøet, antas her uavhengig av genetisk variasjon.,
Den genetiske variansen av dominans er felles for to barn av samme foreldre, fordi de deler dominans effekter når du er på et gitt locus de har fått det samme gener fra begge foreldrene, noe som skjer med dem på én av fire loci. De gjør ikke dele disse effektene med sine foreldre, slik at de blir mer «lignende» til hverandre enn de er til deres foreldre, til tross for at de deler så mange gener med hverandre. Sikkert, kan en forelder, ved en tilfeldighet, har fått det samme to alleler som ett av sine barn på et gitt locus., Denne sjansen avhenger av frekvensen av alleler i befolkningen. Vi ser at graden av sammenfall mellom et barn og hans eller hennes foreldre for en gitt egenskap er en offentlig eiendom.
Mer generelt, alle komponenter av genetisk varians (Vga+Vgd+Vgi) kan variere mellom bestander. I avlslinjene, en vanlig sak er innavl, som eksos genetisk varians: som et resultat, en egenskap kan ha samme gjennomsnitt i to populasjoner, men har en annen genetisk variasjon. Muligheten for en oppdretter å velge en egenskap er derfor et kjennetegn på linje, ikke for den egenskap.,
forholdet mellom foreldre og barn er målt ved arvbarhet h2. Det er definert som forholdet mellom additive genetiske variansen til total varians :
h2 = Vga/Vt

(2) Korrelasjon på 0, hvor størrelsen av barn er uavhengig av at foreldre. En verdifull gjennomsnitt forelder har produsert et barn av likegyldig verdi, innen distribusjon av barn. Koordinatene til punktene ligger på en sirkel; forventningen av verdien ligger på en linje parallell til foreldrenes aksen (d)., (3) Middels foreldre-barn-forholdet mellom 0 og 1: koordinatene til punktene ligger på en ellipse. Forventning om deres verdi ikke lenger ligger på halverer koordinere akser (som også er den store aksen av fordelingen ellipse), men på line r regresjon. I gjennomsnitt forelder til verdi har produsert et barn av verdi b (mellom 0 og a) med en viss variasjon. Det er kjent at er tilfellet (3) er en mellomting mellom tilfeller (1) og (2)
Det er vist under ulike forutsetninger i Figur 5., Arvbarhet er evnen til å avgjøre om en funksjon kan velges. Figur 5-3 viser at en oppdretter vil være i stand til, innenfor en krysset generasjon, for å flytte gjennomsnitt av karakteren fra verdien 0 til verdi b hvis han velger individer med en verdi. Vi kan vise at b = h2a. For å velge en egenskap, det er nyttig for å øke arvbarhet ratio ved å standardisere miljøet (og dermed redusere miljømessige varians), og ved å plassere seg selv i de miljømessige forhold som fremmer forekomsten av trekk., Genetiker Waddington har vist at i naturen, en endring av miljøet kan dermed avsløre nye egenskaper som vil bli valgt under utvikling.
Referanser og noter
Cover bildet. Sneglehus Cepaea nemoralis & Cepaea hortensis. André Künzelmann, UFZ.,
Denne formelen kan skrives på en mer konvensjonell måte som: \pi =\frac{n(n-1))}{2L}\sum _{i=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
å Være et eksempel på personer som har studert trekk ble målt, avvik kan defineres som differansen mellom gjennomsnitt av kvadratet av verdier og torget gjennomsnittet av verdiene. Dette tiltaket, som alltid er positiv, indikerer spredningen av enkeltpersoner.
Dette er arvbarhet i streng forstand av begrepet. Arvbarhet i bred forstand er forholdet mellom summen av genetiske avvik til total varians., Denne formelen gir arvbarhet med begge foreldre (den «gjennomsnittlige mor»); anslaget med bare én forelder ville gi h2/2.
De Miljømessige Oppslagsverk for Miljøet av Association des Encyclopédies de l Miljøet et de l’Énergie (www.a3e.fr), kontraktsmessig knyttet til Universitetet i Grenoble Alpes og Grenoble INP, og sponset av det franske vitenskapsakademiet.
Hvis du sitere denne artikkelen:VEUILLE Michel (2021), Genetisk polymorphism og variasjon, Encyclopedia av Miljøet, url : https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
artiklene i Leksikonet av Miljøet er gjort tilgjengelig under vilkårene i Creative Commons BY-NC-SA-lisensen, som gir gjengivelse underlagt: siterer kilde, for ikke å gjøre kommersiell bruk av dem, deling identiske første forhold, reprodusering på hver bruk eller distribusjon omtale av denne Creative Commons BY-NC-SA-lisensen.
Legg igjen en kommentar