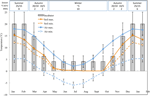
figuur 1. Maandelijkse maximale en minimale lucht-en bodemtemperatuur in de Australische Alpen ( gemiddelde ± STDEV). Luchttemperaturen zijn gemiddelden uit de BOM-database, bodemtemperaturen werden gemeten met iButtons (zie methoden). Bar geeft de temperatuur van de experimentele incubator aan (gemiddelde ± STDEV)., De equivalente tijd van het jaar (seizoen), de incubatortemperatuur (°C, 12/12 uur, licht/donker) en de duur van de behandeling (weken) zoals aangegeven in het bovenpaneel.
Met het oog op het blootleggen van de temperatuur dicht bij die zaden ervaring post-verspreiding, dagelijkse maximum en minimum bodemtemperatuur gegevens werden verzameld binnen het studiegebied (Seaman ‘ s Hut; 36.27°S en 148.17°E, 2030 m een.s.l.) met behulp van ibutton dataloggers (n = 10, Embedded Data Systems, USA) geplaatst 4 cm onder het bodemoppervlak in de basis van de vegetatie van 17 januari tot en met 17 December 2012., We plaatsten loggers op 4 cm om verstoring van het oppervlak te voorkomen, maar nog steeds representatief zijn voor de omstandigheden waaraan zaden worden blootgesteld in de bodem; deze diepte is intermediair aan wat is gebruikt in eerdere Alpine bodem temperatuur monitoring oefeningen (Scherrer en Körner, 2011; Pauli et al., 2015). Zoals vermoed, daalde de gemiddelde bodemtemperatuur in de winter niet onder het vriespunt (figuur 1). In de zomer is het bekend dat de temperatuur van de bodem onder vegetatie de omgevingstemperatuur kan volgen, terwijl temperaturen van kale grond de omgevingstemperatuur met maximaal 15°C zullen overschrijden (Soil Conservation Service unpublished records, 1960-1970).,
Zaadinzameling en kieming
rijpe zaden van 54 soorten uit 16 families en 37 geslachten werden verzameld tussen januari en April 2009, 2010 en 2011 (zie Tabel 1 voor volledige namen en autoriteiten. Vouchers werden ingediend bij het Australian National Herbarium, Canberra). In totaal vertegenwoordigde de soort meer dan een kwart van de Australische angiosperm flora gevonden in alpine gebieden (Costin et al., 2000, zie Tabel 1), hoewel veel van deze soorten zich ook tot onder de boomgrens uitstrekken., De levensvatbaarheid van alle collecties werd geschat vóór het zaaien in experimentele kiemomstandigheden met behulp van de tetrazoliumchloride (TZ) kleuring techniek (International Seed Testing Association, 2003). Voor meer informatie over het verzamelen en verwerken, zie Bijlage A In aanvullend materiaal.
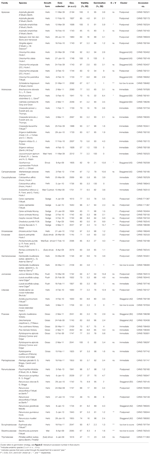
Tabel 1. Studie soort, familie en autoriteit, groeivorm, en collectiedetails, percentage zaad levensvatbaarheid (TZ-geschat, ± S. E.), uiteindelijke percentage kieming bereikt (± S. E.,), statistisch verschil tussen levensvatbaarheid en kieming (G < V, alleen getest in gevallen waarin de kieming minder was dan levensvatbaarheid).
fenologie van zaadkieming werd onderzocht door het nabootsen van temperatuurregimes die alpine zaden in situ ervaren, postdispersie, in een kunstmatig verkorte progressie van seizoenen. Zaden werden ingeslibd gedurende het experiment dus resultaten wijzen op potentieel voor kieming wanneer water niet beperkt., Lucht – en bodemtemperatuurgegevens (figuur 1) werden gebruikt om de temperatuurregimes van incubatoren te sturen en te valideren, hoewel de logistieke contrasten van het werken met incubatoren de integratie van de fluctuaties die inherent zijn aan natuurlijke omstandigheden onmogelijk maakten. Bij de kiemtest werden acht replica ‘ s van 25 zaden per verzameling gebruikt, gezaaid in plastic petrischaaltjes met een diameter van 9 cm met 1% gewoon wateragar. Petrischalen werden verzegeld met Parafilm om agar uitdroging te voorkomen, voordat ze in kiemincubatoren werden geplaatst (Thermoline Scientific, Melbourne, NSW, Australië)., De helft van de schotels was verpakt in aluminiumfolie om licht uit te sluiten en een indicatie te geven van de lichtvereisten voor kieming. Elk van de vier replicaten per lichtbehandeling per collectie werd op een andere plank binnen dezelfde incubator geplaatst en de locatie binnen die plank werd wekelijks opnieuw gerangschikt. Alle replicaten van elke verzameling werden verplaatst door de volgende gesimuleerde thermoperioden: 8 weken bij 25/15°c (dag/nacht temperatuur) → 2 weken bij 20/10 ° c → 2 weken bij 10/5 ° c → 10 weken bij constante 4-5°C → 2 weken bij 10/5 ° c → 2 weken bij 20/10°C → 6 weken bij 25/15°C., Eén cyclus van het hele experiment bootste dus temperaturen na die deden denken aan zomer, vroege herfst, late herfst, winter, vroege lente, late lente en zomer, en duurde in totaal 32 weken. Het verkorte jaar werd redelijk geacht omdat de toetredingen in het algemeen de verdere ontkieming in een bepaalde behandeling hadden stopgezet alvorens naar het volgende “seizoen”te worden overgeheveld., Fluorescentielampen (> 90 µmol m−2 s−1 in het midden van elke plank) zorgden voor een licht/donker fotoperiode van 12/12 uur en in de lente, zomer en herfst viel het licht samen met de warme temperatuurperiode van de dag. Kieming, gedefinieerd als radicle ontstaan met meer dan 1 mm, werd elke 7 dagen gescoord, en gekiemde zaden en zaailingen werden vervolgens verwijderd uit de gerechten. De kieming van folie omwikkelde platen werd als hierboven beoordeeld, maar onder zeer weinig groen licht.,
na één experimentele cyclus (32 weken) werden de meeste collecties beëindigd (zie aanvullende informatie voor uitzonderingen), nadat op dat moment gedurende ten minste 2 weken geen kieming was vertoond. Na beëindiging werden alle resterende intacte zaden ontleed met een scalpel onder een microscoop. Zaden met een stevig, vers endosperm en embryo werden levensvatbaar geacht en zaden die leeg waren van een embryo werden van het totaal afgetrokken bij de berekening van het ontkiemingspercentage voor elke verzameling., We berekenden een index van de Lichtbehoefte voor kieming voor elke soort als een fractie van kieming in de donkere behandeling op het moment dat cumulatieve kiempercentielen van 50 en 95% werden bereikt in de afwisselende licht/donker behandeling (zie hieronder).
ecologische correlaten van de Kiemstrategie
we hebben op basis van gepubliceerde gegevens een database samengesteld van de functionele eigenschappen van soorten en geografische metingen om de correlaties met de kiemstrategie te beoordelen., De databank omvatte de volgende continue kenmerken: gemiddelde zaadmassa, lichte behoefte aan kieming (zie hierboven), specifieke bladoppervlakte (verse oppervlakte/drooggewicht), individuele bladoppervlakte, individueel bladdrooggewicht en lengte bij rijpheid. De minimale, gemiddelde en maximale collectieverhogingen voor de soort werden bepaald op basis van alle herbariumgegevens in het Australische virtuele Herbarium, en de hoogte waarop de specifieke zaadverzamelingen die in het onderzoek werden gebruikt, werden ook opgenomen als continue kenmerken. De groeivorm werd beoordeeld als een van de drie categorieën: graminoïde, kruid of struik., We hebben ook een binaire eigenschap opgenomen: of de zaden endospermisch waren of niet.
de trait-gegevens werden verzameld uit verschillende bronnen, waaronder het Australische virtuele Herbarium (http://avh.chah.org.au/), Het Australian National Herbarium specimen information register (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Alpine Flora (Costin et al., 2000), The New South Wales Flora (Harden, 1990), and the comparative internal morphology of seeds (Martin, 1946). De zaadmassa en het specifieke bladoppervlak werden verkregen uit individuele veldverzamelingen die bij de kiemproeven werden gebruikt., Zaadmassa werd verkregen door het wegen van vier partijen van 10 zaden. SLA werd bepaald op basis van vijf volledig geëxpandeerde bladeren van afzonderlijke individuen per soort. Deze werden verzameld in het veld, gescand op een flatbed scanner, gedroogd bij 60°C tot constant gewicht en gewogen tot op de dichtstbijzijnde 0.000 g. waar onvoldoende zaad beschikbaar was, of specifieke bladoppervlakte niet beschikbaar was, werden gegevens verkregen uit andere anbg collecties van dezelfde soort of uit andere gegevensbronnen (S. Venn, niet-gepubliceerde gegevens)., Of een zaadje wel of niet endospermisch was, werd bepaald aan de hand van tekeningen van het zaadje in de registers van de ANGB seed bank in overleg met het personeel van de ANBG seed bank. De volledige database en bronnen zijn beschikbaar op aanvraag van de auteurs.
Data-analyse
Kiempatronen werden visueel beoordeeld met behulp van een clusteranalyse in PC-ORD voor Windows, versie 6 (MJM Software Design, Gleneden Beach, OR, USA). De analyse maakte gebruik van een Euclidische afstandsmatrix en een dichtsbijzijnde buuralgoritme., Gegevenspunten voor de clusteranalyse waren de tijd (weken) tot specifieke cumulatieve kiempercentielen (t20, t40, t50, t60, t80 en t95%) voor elke verzameling (gemiddeld over de 4 schotels). Zo zou een verzameling die vroeg in het experiment ontkiemde korte tijden tot percentielen tot t80% bijvoorbeeld hebben, terwijl een verzameling die de ontkieming uitstelde tot de temperaturen na de gesimuleerde winter stegen lange tijden tot deze percentielen zou hebben., Complexere ontkiemingspatronen, zoals langzame, gestage ontkieming of uitbarstingen van ontkieming, konden ook worden gedetecteerd, en zouden worden aangegeven door gelijkmatig verdeelde tijden of tijden geclusterd aan het begin en het einde van het experiment, respectievelijk. Deze methode werd gebruikt om een objectieve beoordeling van het kiempatroon op te leggen, onafhankelijk van de temperatuur op zich.,
voor de replicaatplaten van elke verzameling werd een eenrichtingsanalyse van de variantie (ANOVA) uitgevoerd om te beoordelen of er een significant verschil was tussen het percentage levensvatbaarheid (zoals bepaald door de TZ-test) en het uiteindelijke percentage kieming. Verdere algemene lineaire modellen werden gebruikt om te beoordelen of de clusters als gevolg van de clusteranalyse verschilden in levensvatbaarheid of uiteindelijke kiempercentage.
associaties tussen ecologische eigenschappen en clusters van kiemstrategieën werden op twee manieren getest., Ten eerste hebben we getest op gemiddelde verschillen tussen clusters met behulp van ANOVA met cluster als een vast effect en ecologische correlaten als de responsvariabelen, of met behulp van gerepliceerde G-tests voor endospermie en groeivorm. Ten tweede, omdat de kieming en andere functionele eigenschappen hoogst waarschijnlijk door evolutionaire geschiedenis zullen worden beà nvloed, beoordeelden wij deze correlaties gebruikend fylogenetische regressies. Een fylogenie werd geconstrueerd voor de studie soort met behulp van Fylocom Fylomatic en de bladj verpakking in R (Webb et al., 2008; R Core Team, 2013)., We beoordeelden de aanwezigheid van fylogenetische structuur in variabelen met behulp van de phylosig of phylo.d (discrete of binaire variabelen) functie van het Fytoolenpakket in R (Revell, 2012). Fylogenetische regressies werden uitgevoerd met behulp van de pgls-functie in Caper (Orme et al., 2013). Analyses werden uitgevoerd op een gestructureerde manier die de clustering weerspiegelde in PC-Ord (zie hieronder)., In eerste instantie hebben we vier kiemstrategieën opgenomen (uitgestelde, onmiddellijke en gespreide categorieën binnen elk van de voorgaande), dan drie (Alle gespreide soorten op een hoop gooien en vergelijken met uitgestelde en intermediaire) en tenslotte hebben we de verschillen beoordeeld tussen de twee breedste strategieën, uitgesteld en intermediair.
resultaten
levensvatbaarheid van het zaad vergeleken met de Eindkieming
over het geheel genomen waren de levensvatbaarheid van het zaad en het eindkiemingspercentage hoog. De gemiddelde TZ-geschatte levensvatbaarheid voor alle collecties was 80 ± 1.,7%, waarbij meer dan twee derde van de collecties meer dan 75% levensvatbaar is (Tabel 1). Er was geen verschil in de kwaliteit van de ingezamelde zaden over de drie jaar (de gemiddelde levensvatbaarheid in 2009, 2010 en 2011 was respectievelijk 79 ± 2,8, 80 ± 2,7 en 81 ± 3,7%), wat erop wijst dat de banking van de in 2009 ingezamelde zaden hun levensvatbaarheid niet verminderde.
kiemkracht die groter was dan of niet significant lager was dan de TZ-geschatte levensvatbaarheid werd bereikt voor 45 van de 54 soorten (One-Way ANOVA: P>0,05, Tabel 1)., Van de overige negen soorten was de uiteindelijke kieming van vier soorten zeer laag (< 5%), ondanks een goede levensvatbaarheid (ca. 55-94%), en dus konden we GEEN conclusies trekken over de kiemstrategieën van deze soorten. Het niet ontkiemen kan erop wijzen dat onze behandelingen de rust niet verlichtten of dat de zaden niet volledig rijp waren bij het verzamelen. Uit de Cut-tests aan het einde van het experiment bleek dat in alle gevallen het merendeel van de niet-gekiemde zaden een gezond, ingeslibd embryo bevatte., Interessant is dat 34 soorten een grotere kieming bereikten dan de TZ-geschatte levensvatbaarheid (Tabel 1), wat suggereert dat de TZ-test de levensvatbaarheid van zaad doorgaans onderschat.
Kiemclusters
clusteranalyse toonde twee belangrijke clusters aan (A en B), met elk een substructuur (Figuur 2). Voor de meeste soorten (n = 21 soorten uit 10 families) werd de ontkieming uitgesteld tot tijdens een periode van wintertemperaturen (5°c dag/nacht) of tot de temperaturen na de winter werden verhoogd tot 10/5°C (A1 en A1:winter, figuren 2, 3A)., Alternatief, kieming begon onmiddellijk bij 25/15°c (n = 17 soorten, zes families, gezien in cluster B1, figuren 2, 3B). Voor de meeste van deze soorten was het kiempercentage snel; 50% van de totale ontkieming (t50) vond plaats in minder dan 3 weken (aanvullend figuur 1).
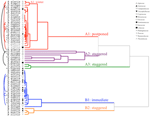
Figuur 2. Clusteranalyse dendrogram gebaseerd op tijd tot cumulatieve kiempercentielen (20, 40, 50, 60, 80, en 95%). Soorten aangeduid met genus initiaal en eerste deel van de soortnaam. Sterretjes markeren collecties die voorafgaand aan het testen werden opgeslagen., Fijne bogen verbinden dubbele collecties om het effect van bankieren op ontkieming te vergelijken. Symbolen duiden verschillende families aan (zie legende). Collecties getoond in grijze tekst zijn die welke werden blootgesteld aan twee cycli van het experiment.
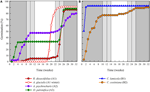
Figuur 3. Cumulatief kiempercentage van één representatieve soort uit elke kiemcluster. (A) cluster A, soorten die de ontkieming uitstellen, (B) cluster B, soorten die onmiddellijk ontkiemden., Merk op dat gespreide kiempatronen binnen de bovenstaande clusters zijn genest. De temperatuurregimes van de Incubator worden weergegeven door arcering (zie Figuur 1).
binnen elk van deze brede klassen waren er echter duidelijke subclusters die de kiemkracht weerspiegelden die in de loop van de tijd werd gespreid, d.w.z. een deel van de partij zaad dat vóór blootstelling aan wintertemperaturen ontkiemde, terwijl de rest na de winter ontkiemde. Trapsgewijze kieming vond plaats bij 13 soorten uit zes families en vond voornamelijk plaats bij een dagtemperatuur boven de 10°C (figuren 3A,B)., Deze gespreide ontkiemmers werden in de clusteranalyse in drie groepen onderverdeeld. Cluster A2 met zeven collecties vertoonde zowel voor als na de koude temperatuurperiode een aanzienlijke ontkieming. Waar de ontkieming stopte voordat de temperatuur veranderde, concludeerden we dat het ontbreken van verdere ontkieming voor de winter niet werd veroorzaakt door de temperatuur (A3). Daarentegen bleek de ontkieming van een groot deel van de A2-groep pas te stoppen wanneer de temperatuur tot 10/5°C werd teruggebracht., Cluster B2 bevatte nog eens vier soorten die in het begin van het experiment een aanzienlijke ontkieming vertoonden en vervolgens een relatief klein deel van hun totale ontkieming bereikten nadat de temperaturen weer boven de 5°C waren gekomen (figuur 3B).
vergeleken met de relatief kleine variatie tussen soorten binnen clusters A1 en B1, vertonen de gespreide clusters ook een veel grotere differentiatie tussen soorten. Individuele krommen voor alle soorten zijn weergegeven in aanvullende figuren 1i-vii. voor meer specifieke details over de kiemstrategie zie Bijlage B bij aanvullend materiaal.,
ecologische correlaten van de Kiemstrategie
van alle ecologische correlaten die we beschouwden, waren alleen de correlaten die direct verband hielden met zaadeigenschappen significant gecorreleerd met de kiemstrategie, ongeacht of we de correlatie beoordeelden op het niveau van twee grote clusters (uitgesteld Versus onmiddellijke kieming), of op het niveau van drie of vier clusters, of of we rekening hielden met fylogenetische structuur of niet (Tabel 2)., Soorten met uitgestelde ontkieming hadden zwaardere zaden en hadden meer kans om endospermisch te zijn dan die van soorten die onmiddellijk ontkiemden (Figuur 4). Soorten met gespreide kieming waren intermediair in zaadmassa en endospermie. Niet alle zaadkenmerken waren echter gecorreleerd met de kiemstrategie: er was geen correlatie tussen de lichte eisen voor de kiemkracht en de strategie. In tegenstelling tot onze verwachtingen, hadden soorten met een hoger specifiek bladgebied (potentieel indicatief voor hogere groeisnelheden) niet meer kans op onmiddellijke ontkieming., Ook soorten met een hogere gemiddelde hoogte of kleinere hoogtegebieden hadden niet meer kans om uitgestelde ontkieming te vertonen.

Tabel 2. Incidentie van fylogenetische structuur in, en correlaten van, kiemstrategie.
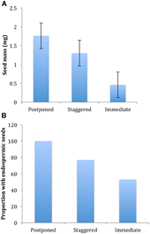
Figuur 4., Verschillen in (a) gemiddelde zaadmassa en (B) aandeel van soorten met endospermische zaden tussen soorten met uitgestelde (cluster A1), gespreide (combinatie van cluster A2, A3 en B2) en directe kieming (cluster B1) strategieën. De verschillen tussen de strategieën zijn significant bij P ≤ 0,05, zie Tabel 2.
discussie
kiemkracht in alpiene habitats werd historisch gezien geacht te zijn gecontroleerd door het milieu, waarbij de wintersneeuw zaden afschermt tegen mogelijke kiemsignalen en slaaptijd overbodig maakt (Billings and Mooney, 1968)., Echter, onze resultaten ondersteunen meer recent bewijs van slapende mechanismen die gemeenschappelijk zijn onder alpine soorten, zowel in Australië (Hoyle et al., 2013; Sommerville et al., 2013) en elders in de wereld (Cavieres and Arroyo, 2000; Schutz, 2002; Shimono and Kudo, 2005; Mondoni et al., 2009; Schwienbacher et al., 2011)., Slaap lijkt een belangrijke rol te spelen bij het beheersen van de in situ timing van ontkieming van veel Australische alpiene soorten, die de snelheid van ontkieming vertragen, uitstellen of vertragen en dus potentieel risico-averse regeneratiestrategieën in een barre en variabele omgeving opleveren. Misschien het meest opvallende van onze kiemresultaten was het aandeel van de soorten waarvoor de kiemstrategie varieerde binnen een zaadverzameling voor een bepaalde soort, wat suggereert dat zowel slapende als niet-slapende kenmerken werden tentoongesteld binnen dezelfde zaadverzameling van deze soorten., In het bijzonder werden kiemstrategieën over het algemeen sterk geconserveerd binnen families en ook gecorreleerd met elementen van zaadanatomie: massa en endospermie. Kiemstrategieën waren echter onafhankelijk van andere ecologisch significante functionele eigenschappen van de volwassen plant.
variatie in Kiemstrategie
zaden van bijna de helft van de onderzochte soorten bleken niet in staat te ontkiemen totdat ze werden blootgesteld aan een koele, natte periode (constante 5°C), wat erop wijst dat koude stratificatie een fysiologisch slaapmechanisme verlichtte., Het uitstellen van de ontkieming tot het volgende voorjaar kan zaailingen in staat stellen om te voorkomen dat de oprichting over of voor de strenge winter, terwijl ook het optimaliseren van de korte komende groeiseizoen. De opportunistische ontkieming van de onmiddellijk ontkiemde soort kan daarentegen een selectief voordeel opleveren wanneer het risico op sterfte van de zaailing in de winter laag is, doordat planten eerder in het volgende voorjaar of op grotere schaal kunnen bloeien (Donohue, 2002).,
tot op heden is er weinig gepubliceerd bewijs van intra-specifieke variabiliteit in de slaapmechanismen binnen alpiene zaadverzamelingen, wat de hier waargenomen gespreide kiemstrategieën zou verklaren. De in de loop van de tijd gespreide kiemkracht kan ook worden verklaard door de wisselende mate van zaadrijpheid bij individuen in de populatie op het moment van de verzameling, en/of kan een dimorfe strategie binnen een enkele plant weerspiegelen die samenhangt met de positie op de plant of het tijdstip van ontwikkeling.,
evolutionaire bet-hedging wordt vaak opgeroepen om de gediversifieerde strategie in zaadkiemkarakteristieken te verklaren (Simons, 2011), zoals die wordt aangetoond door de collecties met gespreide kieming. Als een gediversifieerde strategie echter zowel leidt tot een verhoogde gemiddelde fitness als tot een vermindering van de variantie (bijvoorbeeld door de concurrentie tussen broers en zussen te verminderen), dan is dit geen bet-hedging. Evenzo, als de schijnbare diversificatie weerspiegelt verschillen tussen, niet binnen, individuen dan is het niet bet-hedging (Starrfelt and Kokko, 2012)., Onderzoek naar de verspreiding van alpiene zaailingen en de tolerantie van vorst en sneeuwsmelt, in combinatie met onderzoek naar determinanten van variatie in de kiemstrategie binnen soorten, kan helpen om de kiemfenologie van de soorten die een gespreide strategie vertonen, te verklaren en kan aangeven of zij waarschijnlijk bet-hedging strategieën vertegenwoordigen.,
ecologische correlaten van Kiemstrategie
ons onderzoek naar de relatie tussen kiemstrategie en andere ecologisch belangrijke eigenschappen toonde aan dat kiemeigenschappen gecorreleerd waren met andere zaadeigenschappen maar niet met vegetatieve eigenschappen. Sommerville et al. (2013) vond dat Australische alpenkruiden met niet-endospermische zaden meer kans hadden om niet-slapend te zijn bij verspreiding. Ons werk ondersteunt die bevinding en geeft verder aan dat kleinere zaden meer kans hebben om tot de directe ontkiemers te behoren. Hoewel Sommerville et al., (2013) gaf niet aan waarom endospermie geassocieerd kan worden met rustperiode, wij stellen voor dat het plaatsen van reserves in cotyledons vroege groeisnelheid en vestiging in onmiddellijke kiemende soorten kan verbeteren. Onze resultaten wijzen niet op een correlatie tussen de Lichtbehoefte en de rustperiode, hoewel de Lichtbehoefte per soort verschilt. Er zijn een verscheidenheid aan andere zaadeigenschappen die het risico van uitsterven in variabele omgevingen bufferen, bijvoorbeeld zaadverspreidingsmechanismen en zaadlevensduur in de bodemzaadbank (zie Tielborger et al., 2012 voor een overzicht)., In dit stadium kunnen we niet zeggen of die eigenschappen correlaties zouden vertonen met de kiemstrategie of niet.
we vroegen verder of kiemstrategie zou worden gecorreleerd met ecologisch belangrijke niet-reproductieve eigenschappen, zoals specifieke bladoppervlakte of planthoogte (Westoby, 1998; Wright et al., 2004). In het bijzonder stelden we dat de onmiddellijke kiemende soorten karakteristieke waarden zouden kunnen vertonen die wijzen op een snellere groei (gebruikmakend van SLA als een proxy) zoals nodig zou kunnen zijn voor de vestiging in de korte periode tussen verspreiding en het begin van de winter., We vonden echter geen dergelijke correlaties. Of fysiologische en groeikenmerken van zaailingen in de vroege stadia van de vestiging andere patronen zouden kunnen vertonen dan volwassen vegetatieve kenmerken is een vraag die het overwegen waard is, maar buiten de reikwijdte van het huidige werk.
ten slotte hebben we onderzocht of soorten uit hogere hoogtes waar de groeiseizoenen over het algemeen korter zijn, meer kans zouden hebben om in slaap te vallen, maar er was geen verband tussen hoogteverschil, collectieverhogingen, of de gemiddelde hoogte van de verzameling voor de soort als geheel en kiemstrategie., Samen geven deze resultaten aan dat de ontwikkeling van de kiemstrategie weliswaar nauw verbonden is met ecologisch belangrijke zaadeigenschappen, maar dat deze een grotendeels onafhankelijke as vormt ten opzichte van vegetatieve of verdelingseigenschappen.
implicaties voor alpiene plantengemeenschappen in een veranderend klimaat
Australische alpiene ecosystemen, net als alpiene gebieden over de hele wereld, worden bedreigd door klimaatverandering in combinatie met veranderingen in de brandfrequentie, landgebruikspatronen, instroom van invasieve soorten en de effecten van toegenomen menselijk bezoek., Opwarming in verband met klimaatverandering vindt sneller plaats boven de boomgrens dan op lagere hoogtes, en naar verwachting zullen alpiene gebieden in de toekomst bovengemiddeld opwarmen (Kullman, 2004). De capaciteit voor verdere regeneratie via zaad onder nieuwe omstandigheden zal waarschijnlijk een belangrijke rol spelen bij de reactie van alpiene planten op klimaatveranderingen en daarmee samenhangende veranderingen. Bovendien zullen gewijzigde ordeningsregimes waarschijnlijk een grotere rol van herstel en herstel in de regio vereisen., De inzichten die hier worden verschaft zijn daarom van cruciaal belang voor het behoud en het beheer van alpine systemen onder verandering; ze kunnen helpen om onze voorspellingen over hoe studie soorten kunnen worden beïnvloed, vormen de basis van zaad propagatieplannen voor deze soorten, en kunnen worden gebruikt om toekomstig onderzoek naar alpine zaad kieming strategieën met identificeerbare behoud en beheer resultaten.,
veranderingen in temperatuur zullen waarschijnlijk de rusttoestand van zaden zowel pre – als post-dispersie beïnvloeden, waardoor kiemstrategieën worden gewijzigd en / of verstoord en dit resulteert in verschuivingen in de huidige kiemfenologie (Hoyle et al., 2008b). Voor alpiene planten kan de opwarming van het klimaat leiden tot een verschuiving van het ontstaan van het voorjaar naar het ontstaan van het najaar, voornamelijk gedreven door veranderingen in de rusttoestand van het zaad en resulterend in belangrijke gevolgen voor soorten die momenteel zijn aangepast aan het ontstaan in het voorjaar (Mondoni et al., 2012)., Verder onderzoek naar hoe veranderingen in temperaturen die zaden ervaren na de verspreiding, verminderde duur van de sneeuwbedekking en verhoogde frequentie van extreme temperatuur gebeurtenissen van invloed zullen zijn op de kiemkracht zijn nodig om te voorspellen hoe soorten met rusttijd zullen reageren op klimaatverandering. Voor alpensoorten die in de herfst, voorafgaand aan vorst en sneeuwval, ontkiemen, roepen onze resultaten de vraag op Hoe zaailingen van deze soorten zullen omgaan met de voorspelde vermindering van neerslag en verhoogde frequentie van A-seizoensgebonden vorst gebeurtenissen.,
ten slotte roepen onze resultaten belangrijke vragen op met betrekking tot variatie binnen soorten, met name in verband met het voorspellen van de impact van klimaatverandering of het selecteren van welke zaden te gebruiken voor herstel. Kiemstrategieën kunnen variëren binnen soorten afhankelijk van populatie, bloeitijd, zaadmassa, ouderlijke investering, klimaat, hoogte, en/of maternale omgeving en deze variatie kan genetische, epigenetische en / of omgevingsfactoren weerspiegelen (Cochrane et al., 2015)., Toenemende hoogte is verantwoordelijk voor variatie in kiembaarheid en / of rusttoestand van verschillende alpiene soorten (Billings and Mooney, 1968; Dorne, 1981; Cavières and Arroyo, 2000) maar zie (Hoyle et al., 2014). Verminderde ruststatus kan het gevolg zijn van vroegtijdige beëindiging van zaadontwikkeling als gevolg van milieustress (Steadman et al., 2006; Hoyle et al., 2008b)., Onderzoek naar variatie in kiemstrategieën binnen soorten kan het potentieel voor soorten om te gaan met, en zich aan te passen aan, veranderingen in het klimaat aan het licht brengen, waardoor het een belangrijke rol speelt bij het bepalen van toekomstige overleving en verspreiding van soorten en belangrijke inzichten biedt voor beheer en behoud.
conclusies
onze aanpak heeft een wijdverbreid voorkomen van slaap in de Australische Alpenflora aan het licht gebracht en toont aan dat de kiemstrategieën variëren binnen een zaadverzameling en tussen collecties., Verder, hebben wij aangetoond dat terwijl de slapende strategie phylogenetically wordt behouden en met zaadmassa en endospermy gecorreleerd, het grotendeels onafhankelijk van vegetatieve trekken en waaierkenmerken is. Hoewel men niet direct uit onze resultaten kan afleiden hoe het klimaat en de daarmee samenhangende veranderingen de Alpenflora zullen beïnvloeden, kunnen deze resultaten de basis vormen voor het ontwerp van voortplantingsplannen en verdere experimenten op de impact van veranderende klimaatregimes., In het bijzonder pleiten we voor verder onderzoek naar de oorzaken van binnen-soorten variatie in kiemstrategie en de rol die dit kan spelen in het vermogen van soorten om voorspelde klimaatverandering te weerstaan.
data Accessibility
volledige kiemkracht data en trait database zal worden ingediend op DRYAD en met de Atlas Of Living Australia.
belangenconflict verklaring
De auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.,
Dankbetuigingen
We erkennen de financiering van de Australian Research Council (LP0991593) en de Australian National Botanic Garden en de vrienden daarvan. Wij danken J. McAuliffe, S. Fethers en S. Lawatsch van de ANBG. We danken ook M. Holloway-Phillips, M. Cardillo en S. Cunningham voor input over eerdere versies van het manuscript en voor analytisch advies en S. Venn voor toegang tot trait data.
aanvullend materiaal
Foley, M. E. (1992). Effect van oplosbare suikers en gibberellic-zuur bij het breken van de slaap van uitgesneden wilde haver (Avena fatua) embryo ‘ s., Weed Sci. 40, 208–214.
Google Scholar
Harden, G. J. (1990). Flora of New South Wales. Sydney, NSW: Royal Botanic Gardens.
Google Scholar
International Seed Testing Association. (2003). Werkbladen voor het testen van Tetrazolium. Bassersdorf: ISTA.
Kullman, L. (2004). Het veranderende gezicht van de Alpenwereld. Klodder. Wijzig Newsl. 57, 12–14.
Google Scholar
Geef een reactie