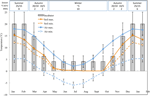
Rysunek 1. Miesięczne dane dotyczące maksymalnej i minimalnej temperatury powietrza i gleby w Alpach Australijskich (średnia ± STDEV). Temperatury powietrza są średnie z bazy danych BOM, temperatury gleby były mierzone za pomocą ibuttonów (patrz metody). Pręty wskazują eksperymentalne temperatury inkubatora (średnia ± STDEV)., Równoważna pora roku (pora roku), reżim temperatury inkubatora (°c, 12/12 h, światło/ciemność) i czas trwania leczenia (tygodnie), jak wskazano na górnym panelu.
w celu odkrycia warunków temperatury zbliżonych do tych, które doświadczają nasion po rozsiewie, dzienne dane dotyczące maksymalnej i minimalnej temperatury gleby zostały zebrane w miejscu badania (Seaman ' s Hut; 36,27°s i 148,17°E, 2030 m n. p. m.) za pomocą rejestratorów danych ibutton (n = 10, Embedded Data Systems, USA) umieszczonych 4 cm pod powierzchnią gleby, na wysokości ok.baza roślinności od 17 stycznia do 17 grudnia 2012., Aby uniknąć zakłóceń na powierzchni, umieściliśmy drwale na wysokości 4 cm, ale nadal są one reprezentatywne dla warunków, na jakie nasiona są narażone w glebie; ta głębokość jest pośrednia do tego, co było stosowane we wcześniejszych ćwiczeniach monitorowania temperatury gleby alpejskiej (Scherrer and Körner, 2011; Pauli et al., 2015). Jak podejrzewano, średnia temperatura gleby w okresie zimowym nie spadła poniżej zera (ryc. 1). Latem temperatura gleby pod roślinnością jest znana z pomiaru temperatury otoczenia, podczas gdy temperatura gołej gleby przekroczy temperaturę otoczenia nawet o 15°C(Soil Conservation Service unpublished records, 1960–1970s).,
zbiór i kiełkowanie nasion
dojrzałe nasiona 54 gatunków z 16 rodzin i 37 rodzajów zostały zebrane między styczniem a kwietniem 2009, 2010 i 2011 (pełne nazwy i władze patrz Tabela 1. Bony zostały złożone w Australian National Herbarium w Canberze). W sumie gatunek ten reprezentował ponad jedną czwartą australijskiej flory angiosperm Phylogeny występującej w regionach alpejskich (Costin et al., 2000, zob. tabela 1), choć wiele z tych gatunków rozciąga się również poniżej linii drzew., Żywotność wszystkich zbiorów oszacowano przed siewem w eksperymentalnych warunkach kiełkowania przy użyciu techniki barwienia chlorkiem tetrazolium (TZ) (International Seed Testing Association, 2003). Więcej informacji na temat zbierania i przetwarzania znajduje się w Dodatku A w materiałach uzupełniających.
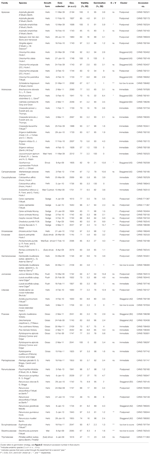
Tabela 1. Zbadaj gatunek, rodzinę i autorytet, formę wzrostu i szczegóły zbiorów, procentową żywotność nasion (TZ-szacowana, ± s. E.), końcowy procent osiągniętego kiełkowania (± s. E.,), statystyczna różnica między żywotnością a kiełkowaniem (G < V, testowana tylko w przypadkach, gdy kiełkowanie było mniejsze niż żywotność).
Fenologię kiełkowania nasion badano poprzez naśladowanie reżimów temperaturowych, które nasiona alpejskie doświadczają in situ, po dyspersji, w sztucznie skróconym przebiegu pór roku. Nasiona zostały wchłonięte przez cały eksperyment, a wyniki wskazują na możliwość kiełkowania, gdy woda nie ogranicza., Dane dotyczące temperatury powietrza i gleby (Rys. 1) zostały wykorzystane do określenia i walidacji reżimów temperaturowych inkubatorów, chociaż logistyczne ograniczenia pracy z inkubatorami wykluczały włączenie wahań właściwych warunkom naturalnym. W badaniach kiełkowania wykorzystano 8 replikatów po 25 nasion na zbiór, wysiewanych na plastikowe płytki Petriego o średnicy 9 cm zawierające 1% zwykłego agaru wodnego. Płytki Petriego zostały zapieczętowane przy użyciu Parafilmu, aby uniknąć wysuszenia agaru, przed umieszczeniem w inkubatorach kiełkowania (Thermoline Scientific, Melbourne, NSW, Australia)., Połowa naczyń została owinięta folią aluminiową, aby wykluczyć światło i zapewnić wskazanie wymagań świetlnych dla kiełkowania. Każda z czterech replikatów na zabieg świetlny na kolekcję została umieszczona na innej półce w tym samym inkubatorze, a lokalizacja w tej półce była zmieniana co tydzień. Wszystkie replikaty każdego zbioru zostały przeniesione przez następujące symulowane termoperiody: 8 tygodni w temperaturze 25/15°C (temperatura dzień/noc) → 2 tygodnie w temperaturze 20/10°C → 2 tygodnie w temperaturze 10/5°C → 10 tygodni w temperaturze stałej 4-5°C → 2 tygodnie w temperaturze 10/5°C → 2 tygodnie w temperaturze 20/10°C → 6 tygodni w temperaturze 25/15°C., Tak więc jeden cykl całego eksperymentu naśladował temperatury przypominające lato, wczesną jesień, późną jesień, zimę, wczesną wiosnę, późną wiosnę i lato i trwał łącznie 32 tygodnie. Skrócony rok został uznany za rozsądny, ponieważ w danym okresie przed przesunięciem na następny „sezon”na ogół zaprzestano dalszego kiełkowania., Światło 12/12 H / ciemny fotoperiod był dostarczany przez świetlówki (> 90 µmol m-2 s-1 na środku każdej półki), a w reżimach temperatury wiosna, lato i jesień światło zbiegło się z ciepłym okresem temperatury w ciągu dnia. Kiełkowanie, określane jako powstawanie rodników o więcej niż 1 mm, było punktowane co 7 dni, a kiełkujące nasiona i sadzonki były następnie usuwane z naczyń. Kiełkowanie płyt owiniętych folią oceniono jak powyżej, ale przy bardzo słabym świetle zielonym.,
po jednym cyklu eksperymentalnym (32 tygodnie) większość zbiorów została przerwana (wyjątki znajdują się w dodatkowych informacjach), wykazując zerowe kiełkowanie przez co najmniej 2 tygodnie w tym czasie. Po terminacji wszystkie pozostałe nienaruszone nasiona zostały rozcięte skalpelem pod mikroskopem. Nasiona z twardym, świeżym bielmem i zarodkiem uznano za żywotne, a nasiona puste z zarodka odejmowano od całkowitej wartości przy obliczaniu procentowego kiełkowania dla każdej kolekcji., Obliczyliśmy wskaźnik zapotrzebowania na światło dla kiełkowania dla każdego gatunku jako ułamek kiełkowania w ciemnym leczeniu w czasie, gdy skumulowane percentyle kiełkowania 50 I 95% zostały osiągnięte w naprzemiennym świetle / ciemnym leczeniu(patrz poniżej).
ekologiczne korelaty strategii kiełkowania
na podstawie opublikowanych danych zebraliśmy bazę cech funkcjonalnych i miar geograficznych gatunków w celu oceny korelacji ze strategią kiełkowania., Baza danych obejmowała następujące ciągłe cechy: średnia masa nasion, zapotrzebowanie na światło do kiełkowania (zob. powyżej), specyficzna powierzchnia liści (świeża powierzchnia/sucha masa), powierzchnia liści indywidualnych, sucha masa liści indywidualnych oraz wysokość w momencie dojrzałości. Minimalne, średnie i maksymalne wysokości zbioru dla gatunku zostały określone na podstawie wszystkich zapisów zielnika w australijskim wirtualnym Zielniku, a wysokość, przy której dokonano określonych zbiorów nasion użytych w badaniu, została również uwzględniona jako cechy ciągłe. Forma wzrostu oceniana była jako jedna z trzech kategorii: graminoid, ziele lub krzew., Uwzględniliśmy również cechę binarną: czy nasiona były bielmo, czy nie.
dane dotyczące cech zostały zebrane z różnych źródeł, w tym australijskiego Wirtualnego zielnika (http://avh.chah.org.au/), australijskiego Krajowego Rejestru okazów zielnika (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Alpine Flora (Costin et al., 2000), The New South Wales Flora (Harden, 1990) oraz the comparative internal morphology of seeds (Martin, 1946). Masę nasion i specyficzną powierzchnię liści uzyskano z pojedynczych zbiorów polowych wykorzystywanych w próbach kiełkowania., Masa nasienna uzyskiwana była przez ważenie czterech partii po 10 nasion. SLA określono na podstawie pięciu w pełni rozszerzonych liści od osobnych osobników na gatunek. Zostały one zebrane w polu, zeskanowane za pomocą skanera płaskiego, wysuszone w temperaturze 60°C do stałej masy i ważone z dokładnością do 0,000 g. W przypadku niewystarczającej ilości nasion lub niedostępności określonej powierzchni liści, dane pozyskano z innych zbiorów ANBG tego samego gatunku lub z innych źródeł danych (S. Venn, dane niepublikowane)., To, czy materiał siewny był bielmo, zostało określone na podstawie rysunków materiału siewnego w rejestrach banku nasion ANGB w porozumieniu z pracownikami banku nasion ANBG. Pełna baza danych i źródła są dostępne na życzenie autorów.
Analiza danych
wzorce kiełkowania zostały ocenione wizualnie i za pomocą analizy klastra w PC-ORD dla Windows, Wersja 6 (MJM Software Design, Gleneden Beach, or, USA). W analizie wykorzystano euklidesową macierz odległości i algorytm łączenia najbliższego sąsiada., Punktami danych dla analizy klastra był czas (tygodnie) do specyficznych skumulowanych percentyli kiełkowania (t20, t40, t50, t60, t80 i t95%) dla każdej kolekcji (uśrednione w 4 naczyniach). Tak więc, zbiór, który kiełkował na początku eksperymentu miałby krótkie czasy do percentyli do t80% na przykład, podczas gdy zbiór, który odroczył kiełkowanie do wzrostu temperatury po symulowanej zimie, miałby długie czasy do tych percentyli., Bardziej złożone wzorce kiełkowania, takie jak powolne, stałe kiełkowanie lub wybuchy kiełkowania, mogą być również wykryte i będą wskazywane przez równomiernie rozmieszczone razy lub czasy zgrupowane odpowiednio na początku i końcu eksperymentu. Metoda ta została zastosowana jako sposób narzucenia obiektywnej oceny wzorca kiełkowania, niezależnej od temperatury per se.,
przeprowadzono jednokierunkową analizę wariancji (ANOVA) dla płyt replikowanych każdego zbioru w celu oceny, czy istnieje znacząca różnica między żywotnością procentową (określoną w teście TZ), a końcowym kiełkowaniem procentowym. Dalsze ogólne modele liniowe zostały wykorzystane do oceny, czy klastry wynikające z analizy klastrów różniły się żywotnością lub procentowym współczynnikiem kiełkowania końcowego.
skojarzenia między cechami ekologicznymi a klastrami strategii kiełkowania zostały zbadane na dwa sposoby., Po pierwsze, testowaliśmy średnie różnice między klastrami za pomocą ANOVA z klastrem jako efektem stałym i korelatami ekologicznymi jako zmiennymi odpowiedzi lub za pomocą replikowanych testów G dla bielmo i formy wzrostu. Po drugie, ponieważ kiełkowanie i inne cechy funkcjonalne są bardzo prawdopodobne, aby mieć wpływ na historię ewolucyjną, oceniliśmy te korelacje za pomocą regresji filogenetycznych. Filogeneza została skonstruowana dla gatunku badawczego przy użyciu Phyllocom Phyllomatic i bladjacku w R (Webb et al., 2008; R Core Team, 2013)., Oceniliśmy obecność struktury filogenetycznej w zmiennych za pomocą phyllosig lub phylo.funkcja D (zmienne dyskretne lub binarne) pakietu Phytools w R (Revell, 2012). Regresje filogenetyczne przeprowadzono wykorzystując funkcję pgls w Caper (Orme et al., 2013). Analizy przeprowadzono w sposób ustrukturyzowany, odzwierciedlając klastry ujawnione w PC-Ord(patrz poniżej)., Początkowo uwzględniliśmy cztery strategie kiełkowania (kategorie odroczone, natychmiastowe i rozłożone w ramach każdej z poprzednich), następnie trzy (sumując wszystkie rozłożone gatunki i porównując do odroczonych i pośrednich) i wreszcie oceniliśmy różnice między dwoma najszerszymi strategiami, odroczone i pośrednie.
wyniki
żywotność nasion w porównaniu do końcowego kiełkowania
ogólnie żywotność nasion i końcowy odsetek kiełkowania były wysokie. Średnia żywotność TZ-szacowana we wszystkich kolekcjach wynosiła 80 ± 1.,7%, przy czym ponad dwie trzecie zbiorów wykazuje ponad 75% rentowności(Tabela 1). Nie było różnicy w jakości zbiorów w ciągu 3 lat (średnia żywotność w latach 2009, 2010 i 2011 wynosiła odpowiednio 79 ± 2,8, 80 ± 2,7 i 81 ± 3,7%), co wskazuje, że bankowość nasion zebranych w 2009 r.nie zmniejszyła ich żywotności.
kiełkowanie, które przekroczyło lub nie było znacząco niższe od TZ-szacowana żywotność została osiągnięta dla 45 z 54 gatunków (jednokierunkowa ANOVA: P>0.05, Tabela 1)., Spośród pozostałych dziewięciu gatunków ostateczna kiełkowanie czterech gatunków było bardzo niskie (< 5%), pomimo dobrej żywotności (ok. 55-94%), a zatem nie byliśmy w stanie wyciągnąć żadnych wniosków na temat strategii kiełkowania tych gatunków. Brak kiełkowania mógł odzwierciedlać albo to, że nasze zabiegi nie złagodziły spoczynku, albo że nasiona nie były w pełni dojrzałe w momencie zbierania. Testy cięcia na końcu eksperymentu wykazały, że we wszystkich przypadkach większość nie kiełkujących nasion zawierała zdrowy, nasycony zarodek., Co ciekawe, 34 gatunki osiągnęły kiełkowanie, które było większe niż szacowana żywotność TZ (Tabela 1), co sugeruje, że test TZ powszechnie niedoceniał żywotność nasion.
klastry kiełkowania
analiza klastrów wykazała dwa główne klastry (A i B), z podbudową w każdym (ryc. 2). W przypadku większości gatunków (n = 21 gatunków z 10 rodzin) kiełkowanie odkładano na okres temperatury zimowej (5°c dzień/noc) lub na okres wzrostu temperatury do 10/5°c po zimie (A1 i A1:zima, Rys. 2, 3A)., Alternatywnie, kiełkowanie rozpoczęło się natychmiast w temperaturze 25/15°C (N = 17 gatunków, sześć rodzin, widzianych w klastrze B1, Rys. 2, 3b). U większości tych gatunków szybkość kiełkowania była szybka; 50% całkowitego kiełkowania (t50) wystąpiło w czasie krótszym niż 3 tygodnie (rysunek dodatkowy 1).
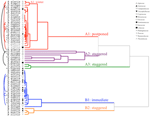
Rysunek 2. Analiza klastrów dendrogram na podstawie czasu do skumulowania percentyli kiełkowania (20, 40, 50, 60, 80, I 95%). Gatunek wskazany przez rodzaj i pierwszą część nazwy gatunkowej. Gwiazdki oznaczają kolekcje, które zostały zablokowane przed testowaniem., Łuki cienkie łączą zbiory duplikatów, aby porównać wpływ bankowości na kiełkowanie. Symbole oznaczają różne rodziny (patrz legenda). Kolekcje pokazane szarym tekstem to te, które zostały wystawione na dwa cykle eksperymentu.
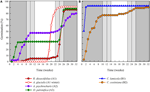
Rysunek 3. Skumulowany procent kiełkowania jednego reprezentatywnego gatunku z każdej gromady kiełkowania. A) Gromada A, gatunki, które opóźniły kiełkowanie, B) Gromada B, gatunki, które natychmiast kiełkowały., Zauważ, że rozłożone wzorce kiełkowania są zagnieżdżone w powyższych klastrach. Systemy temperatury inkubatora są reprezentowane przez cieniowanie (patrz rysunek 1).
w ramach każdej z tych szerokich klas istniały jednak wyraźne podgrupy odzwierciedlające kiełkowanie, które było rozłożone w czasie, tzn. część partii nasion kiełkowała przed wystawieniem na działanie temperatur zimowych, podczas gdy pozostała część kiełkowała po zimie. Kiełkowanie rozłożyste wystąpiło u 13 gatunków z sześciu rodzin i wystąpiło przede wszystkim, gdy temperatura w ciągu dnia przekraczała 10°C (ryc. 3A, B)., Te rozłożone germinatory zostały podzielone na trzy grupy w analizie klastra. Klaster A2 z siedmioma zbiorami wykazywał znaczne kiełkowanie zarówno przed, jak i po okresie zimnej temperatury. Gdy kiełkowanie ustało przed zmianą temperatury, doszliśmy do wniosku, że brak dalszego kiełkowania przed zimą nie był spowodowany temperaturą (A3). W przeciwieństwie do tego, kiełkowanie dużej części Grupy A2 wydawało się zatrzymane dopiero wtedy, gdy temperatura została obniżona do 10/5°C., Klaster B2 zawierał kolejne cztery gatunki, które wykazywały znaczną zdolność kiełkowania na początku eksperymentu i osiągnęły stosunkowo niewielką część całkowitej zdolności kiełkowania po powrocie temperatur powyżej 5°C (ryc. 3b).
w porównaniu ze stosunkowo niewielką zmiennością wśród gatunków w obrębie klastrów A1 i B1, klastry rozłożone wykazują również znacznie większe zróżnicowanie wśród gatunków. Poszczególne krzywe dla wszystkich gatunków przedstawiono na rysunkach uzupełniających 1i-vii. szczegółowe informacje na temat strategii kiełkowania przedstawiono w dodatku B w materiale uzupełniającym.,
ekologiczne korelaty strategii kiełkowania
spośród wszystkich ekologicznych korelatów, które rozważaliśmy, tylko te bezpośrednio związane z cechami nasion były znacząco skorelowane ze strategią kiełkowania niezależnie od tego, czy ocenialiśmy korelację na poziomie dwóch głównych klastrów (przełożone vs.natychmiastowe kiełkowanie), czy na poziomie trzech lub czterech klastrów, czy też uwzględnialiśmy strukturę filogenetyczną czy nie (Tabela 2)., Gatunki z opóźnionym kiełkowaniem miały cięższe nasiona i częściej były bielmo w porównaniu z gatunkami, które kiełkowały natychmiast (ryc. 4). Gatunki o rozłożystym kiełkowaniu były pośrednie w masie nasion i bielmo. Jednak nie wszystkie cechy nasion były skorelowane ze strategią kiełkowania: nie było korelacji między wymaganiami dotyczącymi światła dla kiełkowania nasion a strategią. Wbrew naszym oczekiwaniom, gatunki o większej specyficznej powierzchni liści (potencjalnie wskazujące na wyższe tempo wzrostu) nie wykazywały większego prawdopodobieństwa natychmiastowego kiełkowania., Podobnie gatunki o wyższych średnich wysokościach lub mniejszych wysokościach nie wykazywały większego prawdopodobieństwa odroczenia kiełkowania.

Tabela 2. Występowanie struktury filogenetycznej i korelacje strategii kiełkowania.
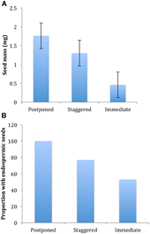
Rysunek 4., Różnice w (A) średniej masie nasion i (B) odsetku gatunków z nasionami endospermicznymi wśród gatunków o strategiach odroczonego (klaster A1), rozłożonego (kombinacja klastrów A2, A3 i B2) i natychmiastowego kiełkowania (klaster B1). Różnice między strategiami są istotne przy P ≤ 0,05, patrz Tabela 2.
dyskusja
kiełkowanie w siedliskach alpejskich było historycznie uważane za kontrolowane ekologicznie, z zimowym śniegiem izolującym nasiona przed potencjalnymi wskazówkami kiełkowania i niepotrzebnym uśpieniem (Billings and Mooney, 1968)., Jednak nasze wyniki potwierdzają nowsze dowody na to, że mechanizmy uśpienia są powszechne wśród gatunków alpejskich zarówno w Australii (Hoyle et al., 2013; Sommerville et al., 2013) i gdzie indziej na świecie (Cavieres and Arroyo, 2000; Schutz, 2002; Shimono and Kudo, 2005; Mondoni et al., 2009; Schwienbacher et al., 2011)., Uśpienie wydaje się odgrywać znaczącą rolę w kontrolowaniu czasu kiełkowania in situ wielu australijskich gatunków alpejskich, działając w celu opóźnienia, opóźnienia lub spowolnienia szybkości kiełkowania, a tym samym potencjalnie nadając strategie regeneracji zapobiegające ryzyku w trudnym i zmiennym środowisku. Być może najbardziej uderzające z naszych wyników kiełkowania był odsetek gatunków, dla których strategia kiełkowania różniła się w ramach zbioru nasion dla danego gatunku, co sugeruje, że zarówno uśpione, jak i nie uśpione cechy były wykazywane w ramach tego samego zbioru nasion tych gatunków., W szczególności strategie kiełkowania były na ogół silnie zachowane w obrębie rodzin, a także skorelowane z elementami anatomii nasion: masą i bielmo. Strategie kiełkowania były jednak niezależne od innych ekologicznie istotnych cech funkcjonalnych dojrzałej rośliny.
zmienność strategii kiełkowania
nasiona prawie połowy badanych gatunków wydawały się niezdolne do kiełkowania, dopóki nie zostały poddane chłodnemu, mokremu okresowi (stała 5°C), co sugeruje, że zimna stratyfikacja łagodzi fizjologiczny mechanizm uśpienia., Odroczenie kiełkowania do następnej wiosny może umożliwić sadzonkom uniknięcie ustanowienia przed lub przed surową zimą, a także optymalizację krótkiego nadchodzącego sezonu wegetacyjnego. Natomiast oportunistyczne kiełkowanie okazywane przez gatunki, które natychmiast kiełkują, może przynieść selektywną korzyść, gdy ryzyko śmiertelności sadzonek zimowych jest niskie, umożliwiając roślinom kwitnienie wcześniej następnej wiosny lub przy większych rozmiarach (Donohue, 2002).,
do tej pory niewiele jest opublikowanych dowodów na zmienność wewnątrzspecyficzną mechanizmów uśpienia w kolekcjach nasion alpejskich, które wyjaśniałyby obserwowane tu strategie kiełkowania. Kiełkowanie rozłożone w czasie może być również wyjaśnione przez różne poziomy dojrzałości nasion wśród osobników w populacji w momencie zbierania i / lub może odzwierciedlać strategię dymorficzną w obrębie jednej rośliny związaną z pozycją na roślinie lub czasem rozwoju.,
ewolucyjne hedgingowanie jest często przywoływane w celu wyjaśnienia zróżnicowanej strategii w cechach kiełkowania nasion (Simons, 2011), takich jak te wykazywane przez zbiory o rozłożonym kiełkowaniu. Jeśli jednak zdywersyfikowana strategia skutkuje zarówno zwiększoną średnią przydatnością, jak i zmniejszeniem wariancji (np. poprzez zmniejszenie konkurencji między rodzeństwem), to nie stanowi zabezpieczenia zakładu. Podobnie, jeśli pozorna dywersyfikacja odzwierciedla różnice między jednostkami, a nie wewnątrz nich, to nie jest to hedging (Starrfelt and Kokko, 2012)., Badania nad wzorami rozprzestrzeniania się sadzonek alpejskich i tolerancją mrozu i roztopów śniegu, w połączeniu z badaniami determinantów zmienności strategii kiełkowania w obrębie gatunków, mogą pomóc wyjaśnić fenologię kiełkowania gatunków wykazujących strategię rozłożoną i mogą wskazać, czy mogą one reprezentować strategie hedgingowe.,
ekologiczne korelacje strategii kiełkowania
nasze badanie związku między strategią kiełkowania a innymi cechami ważnymi ekologicznie wykazało, że cechy kiełkowania były skorelowane z innymi cechami nasion, ale nie z cechami wegetatywnymi. Sommerville et al. (2013) odkrył, że australijskie zioła alpejskie z nie endospermicznymi nasionami były bardziej prawdopodobne, że nie są uśpione w rozproszeniu. Nasza praca wspiera to odkrycie i dodatkowo wskazuje, że mniejsze nasiona są bardziej prawdopodobne, aby znaleźć się wśród bezpośrednich kiełkujących. Chociaż Sommerville et al., (2013) nie zakładał, dlaczego bielmo może być związane z uśpieniem, sugerujemy, że umieszczenie rezerw w liścieniach może poprawić wczesne tempo wzrostu i ustanowienie w natychmiastowych gatunkach kiełkujących. Nasze wyniki nie wykazały żadnej korelacji między zapotrzebowaniem na światło a uśpieniem, chociaż wymagania dotyczące światła różniły się w zależności od gatunku. Istnieje wiele innych cech nasion, które buforują ryzyko wyginięcia w zmiennych środowiskach, na przykład mechanizmy rozpraszania nasion i długowieczność nasion w banku nasion gleby (patrz Tielborger et al., 2012 za recenzję)., Na tym etapie nie możemy powiedzieć, czy te cechy wykazywałyby korelacje ze strategią kiełkowania, czy nie.
zapytaliśmy dalej, czy strategia kiełkowania byłaby skorelowana z ekologicznie ważnymi cechami Nie-reprodukcyjnymi, takimi jak określony obszar liści lub wysokość rośliny (Westoby, 1998; Wright et al., 2004). W szczególności zakładaliśmy, że natychmiastowe gatunki kiełkujące mogą wykazywać wartości cech wskazujące na szybsze tempo wzrostu (przy użyciu SLA jako pośrednika), które mogą być konieczne do ustanowienia w krótkim okresie między rozproszeniem a początkiem zimy., Nie znaleźliśmy jednak takich korelacji. To, czy fizjologiczne i wzrostowe cechy sadzonek na wczesnym etapie zakładania mogą wykazywać inne wzorce niż dorosłe cechy wegetatywne, jest kwestią godną rozważenia, ale wykraczającą poza zakres niniejszej pracy.
na koniec zbadaliśmy, czy gatunki z wyższych wysokości, gdzie okresy wegetacji są na ogół krótsze, mogą wykazywać większe prawdopodobieństwo uśpienia, ale nie było związku między zakresem wzniesień, wzniesieniami kolekcji lub średnim wzniesieniem kolekcji dla gatunku jako całości a strategią kiełkowania., Łącznie wyniki te wskazują, że chociaż ewolucja strategii kiełkowania jest ściśle powiązana z ekologicznie ważnymi cechami nasion, stanowi ona w dużej mierze niezależną oś względem cech wegetatywnych lub dystrybucyjnych.
implikacje dla alpejskich zbiorowisk roślinnych w zmieniającym się klimacie
Australijskie ekosystemy alpejskie, takie jak obszary alpejskie na całym świecie, są zagrożone zmianami klimatu połączonymi ze zmianami częstotliwości pożarów, wzorców użytkowania gruntów, napływu gatunków inwazyjnych i skutków zwiększonej liczby odwiedzin ludzi., Ocieplenie związane ze zmianami klimatycznymi następuje szybciej powyżej linii drzew niż na niższych wysokościach, a obszary alpejskie powinny nadal doświadczać ponadprzeciętnego ocieplenia w przyszłości (Kullman, 2004). Zdolność do dalszej regeneracji poprzez nasiona w nowych warunkach prawdopodobnie odegra znaczącą rolę w reakcji roślin alpejskich na klimat i związane z tym zmiany. Ponadto zmienione systemy zakłócania mogą wymagać zwiększonej roli odbudowy i rehabilitacji w regionie., Przedstawione tutaj spostrzeżenia mają zatem kluczowe znaczenie dla zachowania i zarządzania systemami alpejskimi pod wpływem zmian; mogą pomóc w przewidywaniu, w jaki sposób gatunki badane mogą zostać dotknięte, stanowią podstawę planów rozmnażania nasion dla tych gatunków i mogą być wykorzystane do prowadzenia przyszłych badań nad strategiami kiełkowania nasion alpejskich z możliwymi do zidentyfikowania wynikami ochrony i zarządzania.,
zmiany temperatury mogą wpływać na stan uśpienia nasion zarówno przed, jak i po dyspersji, w konsekwencji zmieniając i / lub zakłócając strategie kiełkowania i powodując zmiany w obecnej fenologii kiełkowania (Hoyle et al., 2008b). W przypadku roślin alpejskich ocieplenie klimatu może prowadzić do przejścia od wiosennego do jesiennego wschodzenia, napędzanego głównie zmianami stanu uśpienia nasion i skutkującego poważnymi konsekwencjami dla gatunków obecnie przystosowanych do wschodzenia wiosną (Mondoni et al., 2012)., Dalsze badania nad tym, w jaki sposób zmiany temperatur, które doświadczają nasion po rozproszeniu, krótszy czas trwania pokrywy śnieżnej i zwiększona częstotliwość ekstremalnych zdarzeń temperaturowych wpłyną na kiełkowanie, są wymagane w celu przewidywania, w jaki sposób gatunki z uśpieniem zareagują na zmiany klimatu. Dla tych alpejskich gatunków, które kiełkują jesienią, przed mrozem i opadami śniegu, nasze wyniki rodzą pytanie, w jaki sposób sadzonki tych gatunków poradzą sobie z przewidywanym spadkiem opadów i zwiększoną częstością sezonowych mrozów.,
wreszcie, nasze wyniki budzą ważne pytania dotyczące zmienności w obrębie gatunków, szczególnie w związku z przewidywaniem wpływu zmian klimatycznych lub wyborem nasion do wykorzystania do odbudowy. Strategie kiełkowania mogą się różnić w obrębie gatunku w zależności od populacji, czasu kwitnienia, masy nasion, inwestycji rodziców, klimatu, wysokości i/lub środowiska matki, a ta zmienność może odzwierciedlać czynniki genetyczne, epigenetyczne i/lub środowiskowe (Cochrane et al., 2015)., Wzrost wysokości odpowiada za różnice w stanie kiełkowania i / lub uśpienia kilku gatunków alpejskich (Billings and Mooney, 1968; Dorne, 1981; Cavieres and Arroyo, 2000), ale zobacz (Hoyle et al., 2014). Obniżony stan uśpienia może wynikać z wczesnego zakończenia rozwoju nasion wynikających z naprężeń środowiskowych (Steadman et al., 2006; Hoyle et al., 2008b)., Badanie zmienności strategii kiełkowania w obrębie gatunków może ujawnić potencjał radzenia sobie ze zmianami klimatu i przystosowywania się do nich, odgrywając w ten sposób ważną rolę w określaniu przyszłego przetrwania i rozmieszczenia gatunków oraz dostarczając ważnych informacji dla zarządzania i ochrony.
wnioski
nasze podejście ujawniło powszechne występowanie uśpienia w australijskiej florze alpejskiej i pokazuje, że strategie kiełkowania różnią się zarówno w kolekcji nasion, jak i między kolekcjami., Ponadto wykazaliśmy, że chociaż strategia uśpienia jest filogenetycznie zachowana i skorelowana z masą nasion i bielmo, jest ona w dużej mierze niezależna od cech wegetatywnych i cech zasięgu. Podczas gdy nie można ekstrapolować bezpośrednio z naszych wyników, w jaki sposób klimat i związane z nim zmiany wpłyną na florę alpejską, wyniki te mogą stanowić podstawę do opracowywania planów rozmnażania i dalszych eksperymentów na temat wpływu zmieniających się reżimów klimatycznych., W szczególności popieramy dalsze badania nad zrozumieniem przyczyn zmienności wewnątrzgatunkowej w strategii kiełkowania i roli, jaką może to odegrać w zdolności gatunków do wytrzymywania przewidywanych zmian klimatycznych.
dostępność danych
Pełna baza danych i cech zostanie umieszczona na DRYAD oraz w Atlasie żywej Australii.
Oświadczenie o konflikcie interesów
autorzy oświadczają, że badanie zostało przeprowadzone w przypadku braku jakichkolwiek relacji handlowych lub finansowych, które mogłyby być interpretowane jako potencjalny konflikt interesów.,
podziękowania
potwierdzamy finansowanie z Australian Research Council (LP0991593) oraz Australian National Botanic Garden i jego przyjaciół. Dziękujemy J. McAuliffe, S. Fethers i S. Lawatsch z ANBG. Dziękujemy również M. Holloway-Phillips, M. Cardillo i S. Cunningham za wkład we wcześniejsze wersje rękopisu i za doradztwo analityczne oraz S. Venn za dostęp do danych cech.
Materiały uzupełniające
Wpływ rozpuszczalnych cukrów i kwasu giberelinowego na rozkład uśpienia wyciętych zarodków owsa Dzikiego (Avena fatua)., Chwast Sci. 40, 208–214.
Google Scholar
Flora Nowej Południowej Walii. Sydney, NSW: Royal Botanic Gardens.
Google Scholar
International Seed Testing Association. (2003). Arkusze robocze dotyczące testów Tetrazolium. Bassersdorf: ISTA.
Kullman, L. (2004). Zmieniające się oblicze alpejskiego świata. Glob. Zmień Newsl. 57, 12–14.
Google Scholar
Dodaj komentarz