
polimorfizm genetyczny to istnienie alternatywnych Stanów DNA, określających zmienność wyższych poziomów integracji organizmu. Istnieją różne rodzaje modyfikacji genomu (mutacji). Najczęściej badane są podstawienia nukleotydów w regionach kodujących i regulacyjnych.
definicja
polimorfizm genetyczny polega na istnieniu w populacji kilku alternatywnych Stanów DNA lub alleli w określonej pozycji w genomie lub locupozycji genu na chromosomie., W genetyce populacyjnej zbiór genów homologicznych (Klasa homologii). Dwa geny są homologiczne, jeśli pasują do mejozy. Liczba mnoga: loci.. Definicja ta ma kilka aspektów:
(1) Po pierwsze, cecha musi być przenoszona przez chromosomy i być przenoszona.
(2) wtedy allele muszą być homologicznymi chromosomami lub mówi się, że dwa geny są homologiczne, jeśli pasują do siebie i wzajemnie wykluczają się z mejozy. dla ich pozycji w genomie, co wyklucza pewne rodzaje zmienności, które zostaną omówione później.,
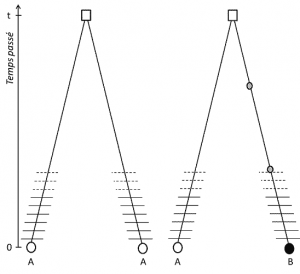
(3) ale ponieważ cecha jest przenoszalna, homologia pozycji zakłada również, że allele są homologiczne przez pochodzenie; jeśli są różne, to dlatego w jednej z linii łączących je z ostatnim wspólnym przodkiem (ryc. 1) wystąpiła mutacja (co najmniej).
(4) Wreszcie polimorfizm genetyczny można zdefiniować w skali najmniejszej jednostki tworzącej DNA: miejsca nukleotydów., W rezultacie każdy wariant nukleotydu może określić polimorfizm wyższych poziomów struktury biologicznej-genu jednostki, białka i fenotypu-które stają się skalami, w których można opisać polimorfizm genetyczny (ryc. 2).
weźmy przykład układu krwi ABO, który odpowiada motywom antygenowymczęść antygenu rozpoznanego przez przeciwciało lub receptor limfocytów. Znany również jako epitop lub wyznacznik antygenowy. Ten sam antygen może mieć kilka epitopów (identycznych lub różnych) i w ten sposób wywoływać zróżnicowaną odpowiedź immunologiczną., zaangażowany w Zgodność podczas transfuzji krwi. Jest to przypadek polimorfizmu genetycznego występującego w populacjach ludzkich, polegającego na transferach białka abo glikozylu, pozwalających na transfer pozostałości zawierających cukry do białek. W przypadku układu ABO, transferaza glikozylowa a i B odpowiednio indukują osobnika do przynależności do grupy A lub B. W przypadku obecności obu glikozyltransferazy osobnik należy do grupy AB. który posiada trzy allele, A, B I O. osobnik może mieć genotypy (AA), (AO); (BB), (BO); (oo) lub (AB)., Jego fenotyp będzie wtedy,,;,; lub, gdzie widzimy, że A i B „dominują” O (tzn. ich ekspresja maskuje to, co o), i że są one dominujące między sobą (heterozygotyczne między A i B ma rozpoznawalną ekspresję,).
polimorfizm można opisać najpierw w skali sekwencji DNA locus kodującego białko. Niektóre polimorfizmy są „synonimiczne”, tzn. nie zmieniają sekwencji aminokwasowej białka; są często najliczniejszymi polimorfizmami. Pozostałe modyfikują aminokwasy i nazywane są polimorfizmami „zastępczymi”., Polimorfizmy zastępcze transferazy GLIKOZYLOWEJ ABO są dwóch typów: polimorfizmy, które zmieniają aminokwas, ale nie jednostkę antygenową (nie zakłócają fenotypu abo osobników); i inne polimorfizmy zastępcze, które determinują fenotyp ABO obserwowalnych cech jednostki..
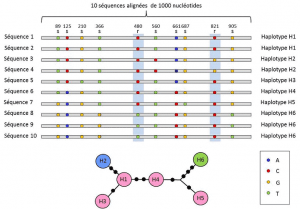
analizy genetyczne uwzględniają pośredni poziom opisu: liniowy układ zmiennych miejsc na locus. Układy te, zwane haplotypamigrupa alleli o różnych loci zlokalizowanych na tym samym chromosomie i zwykle przenoszonych razem. Haplotyp jest utworzony przez skrócenie angielskiego wyrażenia haploid genotyp lub haploid genotyp., Wszystkie geny znajdujące się na tym samym chromosomie, których allele segregują się razem podczas mejozy, stanowią haplotyp. Geny te nazywane są „genetycznie powiązane”, są przydatne w genetyce ewolucyjnej, ponieważ pozwalają znaleźć genealogiczne powiązania między allelami (ryc. 2).
polimorfizmy nukleotydów występują również w regionach międzygatunkowych (poza regionami kodującymi). Niektóre mają wpływ na obszary, w których ekspresja genów jest regulowana i dlatego mają ekspresję fenotypową. Inne nie mają znanego efektu i są określane jako „ciche”.,
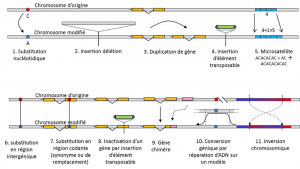
oprócz podstawień nukleotydów, polimorfizm dla określonego typu mutacji, mikrosatelitów, odnosi się do zmian długości często niekodującego fragmentu DNA (ryc. 3) ze względu na zmianę liczby powtórzeń w krótkiej powtarzalnej sekwencji nukleotydów, np. CACACACA lub TGTGTG.
nie każda zmienność genetyczna mieści się w definicji polimorfizmu genetycznego, ponieważ wymaga homologii pozycji alternatyw (ryc. 3)., W związku z tym istnieją powtarzające się sekwencje uzyskane przez powielenie genu, dla którego dwa powtórzenia nie są homologiczne na pozycję. Często nie możemy też mówić o homologii dla elementów transponowalnych, ponieważ na ogół zmieniają one pozycję podczas generacji komórek, mogą się namnażać i atakować Genom. Nie można wówczas zdefiniować homologii dwóch elementów transponowalnych. Z drugiej strony, dwie sekwencje kodujące tego samego locus, z których jedna jest nienaruszona, a druga nieaktywna przez wstawienie transponowalnego elementu, dobrze odpowiadają sytuacji homologicznej., Pewnego dnia prawdopodobnie zastanowimy się nad polimorfizmem modyfikacji tzw.” epigenetycznych ” chromosomów, które są czasami przenoszone przez kilka pokoleń komórek, somatycznych komórek nie rozmnażających się lub soma. Mutacja somatyczna, która wpływa na gen komórki somatycznej, znika wraz z nosicielem. lub kiełkuje gamety. Mutacja germinalna może być przenoszona na potomstwo., np. metylacja. Ich znaczenie w ewolucji populacji pozostaje do oceny., Ograniczenie pojęcia polimorfizmu genetycznego do pewnych kategorii zmienności – zasadniczo substytucji nukleotydowych – wynika z Wielkiej użyteczności tych ostatnich w badaniu historii naturalnych populacji. Nadają się do modelowania matematycznego ewolucji.
słowo polimorfizm oznacza „kilka form”. Jest przeciwny monomorfizmowi, który wskazuje na brak zmienności. W słownictwie biologii opozycja ta od dawna była używana, przed genetyką, do sygnalizowania współistnienia kilku rodzajów osobników w tym samym gatunku, np., polimorfizm kastowy (królowa i robotnice) u owadów społecznych; polimorfizm sezonowy (zmiany futra) u niektórych ssaków arktycznych. Przypadki te nie podlegają polimorfizmowi genetycznemu, nowszemu pojęciu o węższym i bardziej precyzyjnym znaczeniu. Zmiany ciągłe (np. zmienność wielkości) również nie należą do kategorii polimorfizmu, ponieważ nie stanowią odrębnych alternatyw. Jednak locus, które wpływają na rozmiar należą do tej definicji., Poniżej omówimy najczęściej badane odmiany genetyczne, ponieważ są one najważniejsze dla ewolucji, podstawienia nukleotydów, a następnie omówimy zmienność fenotypów.
pomiar
nie wystarczy powiedzieć, że locus jest mniej lub bardziej zmienny bez podania ilościowego oszacowania tego wyroku. Badacz może badać zmienność w różnych skalach. Jeśli interesuje go tylko allel białka, zmierzy on różnorodność alleliczną, symbolizowaną przez „H”. Jeśli interesuje go różnorodność DNA, zmierzy różnorodność nukleotydów, symbolizowaną przez „π”.,
różnorodność alleli H definiowana jest jako prawdopodobieństwo wylosowania dwóch różnych alleli podczas dwóch prób z wymianąpowrót do kolejnego losowania z dostarczeniem żetonów p do urny zawierającej żetony n, czyli wzięcia pierwszego żetonu, odczytania jego wartości, włożenia go z powrotem do urny, wzięcia drugiego żetonu, odczytania jego wartości, włożenia go z powrotem do urny itp., aż do żetonu pth. Oznacza to wybieranie obiektów p spośród n z powtórzeniem (można wybrać ten sam obiekt kilka razy) oraz w kolejności (ważna jest kolejność w jakiej wybieramy obiekty)., Liczba kolejnych losowań ze żetonami wśród n wynosi: n × n × n × … × n = np.. Jeśli nazywamy Pi częstotliwością allelu rangi i, to wykazujemy, że prawdopodobieństwo wybrania dwukrotnie tego samego allelu wynosi F = Σ pi2. Różnorodność alleliczna próbki jest wtedy jej dopełnieniem do 1, tj.:
H = 1 – Σ pi2 (1)
dotyczy to zarówno alleli białkowych, jak i haplotypów. Nazywa się to różnorodnością haplotypową. Można ją również nazwać heterozygotycznością, ponieważ w przypadku diploidalnego locus podaje oczekiwaną częstotliwość heterozygoty.,
różnorodność nukleotydów π jest odpowiednikiem H w skali każdego nukleotydu. Oblicza się ją jako średnią liczbę różnic nukleotydowych między sekwencjami w próbce pobranej dwa na dwa (δij), podzieloną przez długość fragmentu DNA na liczbę nukleotydów (L).
π = średnia (δij) /L (2) (patrz odniesienie)
wartość ta różni się w zależności od gatunku. W kodujących częściach ludzkiego genomu dwa chromosomy losowo wybrane z populacji różnią się średnio o jeden nukleotyd na tysiąc., U muszki owocowej (Drosophila melanogaster) różnica ta wynosi około jednej na sto. Mucha jest więc dziesięć razy bardziej zmienna niż człowiek. Gdy wartości te są związane z wielkością regionów kodujących (około 15 500 genów u Drosophila, co najmniej 22 000 u ludzi), a nawet genomów (140 milionów par zasad na haploidalny Genom U Much, około 3200 milionów lub dwadzieścia razy więcej u ludzi), liczba miejsc polimorficznych jest astronomiczna i powoduje, że nikt nie rodzi się z pokolenia płciowego u żadnego gatunku jest, był i będzie genetycznie identyczny z innym w przeszłości, teraźniejszości lub przyszłości., Siła informacyjna polimorfizmu DNA jest ogromna. W kryminalistyce detektywi mogą zidentyfikować każdego podejrzanego z szesnastu mikrosatelitarnych lokalizacji.
Historia
termin zmienność znajduje się w tytule dwóch pierwszych rozdziałów książki Darwina The Origin of Species (1859). Darwin wprowadził to pojęcie do nauk przyrodniczych w czasie, gdy prawa dziedziczności biologicznej pozostawały tajemnicą i poświęcił mu kolejną ważną książkę, zmienność w zwierzętach i roślinach (1868)., Przekonany, że ewolucja dotyczy małych zmian, które nie mają większego znaczenia dla adaptacji organizmów do ich warunków życia, przywiązywał kluczową wagę do małych zmian ilościowych, co skłoniło jego następców (zwłaszcza Karla Pearsona) do odkrycia biometryzacji pomiaru życia. Odnosi się w bardzo szerokim znaczeniu do ilościowego badania istot żywych.. Jednak ponowne odkrycie praw Mendla w 1900 roku przesunęło zainteresowanie na nieciągłe wariacje.
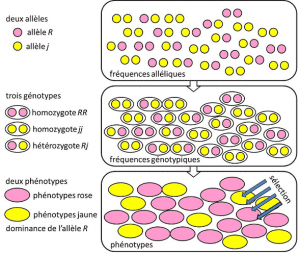
w latach 1908-1930 genetyka populacyjna była zasadniczo dyscypliną teoretyczną dążącą do pogodzenia darwinizmu z dziedzicznością Mendelowską opartą na transmisji pojedynczego genu w dominującym, recesywnym lub powiązanym z chromosomem płci X (lub Y). Odnosi się do Cechy genetycznej z prostym determinizmem, przez parę lub niewielką liczbę par genów. i gdzie prawdopodobieństwo odegrało główną rolę. Genetyka jest intuicyjna., Przewiduje, że nie ma reprodukcji rodzica u jego potomka, ponieważ genotyp tego ostatniego wynika z losowania rozdzielenia alleli przed fuzją dwóch rodzicielskich pół genomów. Zdawano sobie sprawę, że są to częstotliwości alleliczne, przy których allel danego wariantu znajduje się w populacji. Wyrażona jako odsetek lub procent. Suma częstości alleli wszystkich alleli genu w populacji jest więc z definicji równa 1. W genetyce populacyjnej częstość alleli reprezentuje różnorodność genetyczną na poziomie populacji lub gatunku., które są przekazywane między pokoleniami, a nie genotypy czy fenotypy. Częstotliwości te są mniej lub bardziej stabilne z pokolenia na pokolenie i produkują, poza wielokrotnym splątaniem filiacji między osobnikami, te same częstotliwości genotypowegenetyczna struktura populacji. Określone na podstawie częstotliwości allelicznych (rys. 4). Rozkład genotypów populacji, związany z wariancją, jest zatem jedynym przewidywalnym czynnikiem., Około 1930 roku trzej teoretycy Ronald Fisher, JBS Haldane i Sewall Wright pomogli stworzyć pojęcie populacji Mendeliańskiej, której dziedziczność była zgodna z prawami Mendla. . W tym kontekście ewolucja musi wiązać się z trzema poziomami strukturalnymi: genem, osobnikiem i populacją. To trójstronne skojarzenie można podsumować za pomocą następującego wzoru: ewolucja populacji to zmiana częstości alleli (skala genowa, ale także skala grupowa), zmiana zależna od sortowania fenotypów przez selekcję (skala indywidualna)., Empiryczne badania genetyki populacyjnej rozwinęły się, ale przez długi czas były ograniczone przez naszą nieznajomość sposobu działania chromosomów (struktura DNA została zrozumiana w 1953 roku, a jej sekwencjonowanie rozpoczęło się w 1977 roku), i musiały wycofać się z badań nad kilkoma widocznymi polimorfizmami, takimi jak zabarwienie elytry biedronki lub skorupy ślimaków (patrz debata „Wielki ślimak”). Wśród genetyków było wiele dyskusji na temat tego, czy naturalne populacje są na ogół polimorficzne czy monomorficzne, i czy polimorfizm sam w sobie jest korzystny, czy też nie., Badania nad zmiennością skali makromolekularnej musiały poczekać do 1966 (dla białek) i 1983 (dla DNA), zanim odpowiedziały na te pytania.
ilościowe różnice i dziedziczność
Kiedy rozglądamy się wokół siebie, widzimy fizyczne różnice między istotami, niektóre złożone, takie jak rysy twarzy, inne łatwe do zmierzenia, takie jak waga czy wzrost. Zdrowy rozsądek sugeruje, że są one częściowo przenoszone, choć w sposób trudny do określenia. Cechy te są często poligenicznezależność wielu genów. Mówimy o dziedziczności poligenicznej. Cukrzyca jest chorobą poligeniczną.,, co oznacza, że są pod wpływem wielu lokusów. Genetyka rozwojowa i jej zastosowanie do porównywania gatunków (evo-devo, dla „ewolucji rozwoju”) rozwikłają sieć złożonych interakcji, które sprawiają, że podczas rozwoju komórki ciała, wszystkie z identycznym bagażem genetycznym, różnicują się przez epigenezę, aby dać różne tkanki. Nadal nie jest jasne, w jaki sposób złożone postacie są budowane. Mierzalne fenotypy można badać za pomocą biometrii, opartej na właściwościach rozkładów statystycznych., Jedną z wartości o użytecznych właściwościach jest wariancjaw próbie osób, w których mierzono daną cechę, wariancja jest różnicą między średnią kwadratu wartości a kwadratem średniej wartości. Ta miara, która jest zawsze pozytywna, wskazuje na rozproszenie jednostek. . Gdy kilka niezależnych przyczyn określa zmienność linii, ich wariancje są addytywne, a ich suma daje wariancję linii. Jeśli nie są niezależne, do sumy wariancji dodaje się sumę kowariancji., Całkowita wariancja VT cechy fenotypowej jest taka, że :
Vt = Vga+Vgd+Vgd+VGI+Ve
VGA, addytywna wariancja genetyczna, jest sumą wariancji wynikających z każdego locus niezależnie od innych; VGD, Dominująca wariancja genetyczna, wynika z interakcji dwóch alleli tego samego locus; Vgi, wariancja interakcji między locus, wynika z faktu, że locus tego samego osobnika oddziałują; ve, wariancja środowiska, zakłada się tutaj niezależnie od wariancji genetycznej.,
wariancja genetyczna dominacji jest wspólna dla dwojga dzieci tych samych rodziców, ponieważ mają one wspólne efekty dominacji, gdy w danym locus otrzymają te same allele od obojga rodziców, co zdarza się im w co czwartym locus. Nie dzielą się tymi efektami ze swoimi rodzicami, więc są bardziej „podobni” do siebie niż do swoich rodziców, pomimo faktu, że dzielą ze sobą tak wiele genów. Z pewnością rodzic może przypadkiem otrzymać te same dwa allele, co jedno ze swoich dzieci w danym locus., Szansa ta zależy od częstości alleli w populacji. Widzimy, że stopień podobieństwa między dzieckiem a jego rodzicem dla danej cechy jest własnością publiczną.
ogólnie rzecz biorąc, wszystkie składniki wariancji genetycznej (Vga+Vgd+Vgi) mogą się różnić w zależności od populacji. W liniach hodowlanych częstym przypadkiem jest chów wsobny, który wyczerpuje wariancję genetyczną: w rezultacie cecha może mieć taką samą średnią w dwóch populacjach, ale mają inną zmienność genetyczną. Zdolność hodowcy do wyboru cechy jest więc cechą charakterystyczną linii, a nie cechy.,
relacja między rodzicami a dziećmi mierzona jest dziedzicznością h2. Jest on zdefiniowany jako stosunek addytywnej wariancji genetycznej do wariancji całkowitej :
h2 = Vga/Vt

(2) korelacja 0, gdzie wielkość dzieci jest niezależna od wielkości rodziców. Wartościowy Przeciętny rodzic stworzył dziecko o obojętnej wartości, w ramach dystrybucji dzieci. Współrzędne punktów znajdują się na okręgu; oczekiwana ich wartość znajduje się na linii równoległej do osi rodziców (d)., (3) pośrednia korelacja rodzic-dziecko między 0 i 1: współrzędne punktów znajdują się na elipsie. Oczekiwanie ich wartości nie znajduje się już na bisektorze osi współrzędnych (który jest również główną osią elipsy rozkładu), ale na linii regresji r. średni rodzic wartości wytworzył dziecko wartości b (między 0 a a) z pewną wariancją. Należy zauważyć, że przypadek (3) jest pośredni między przypadkami (1) i (2)
jest pokazany w różnych założeniach na rysunku 5., Dziedziczność to zdolność do określenia, czy dana cecha może być wybrana. Rysunek 5-3 pokazuje, że hodowca będzie w stanie, w ramach pokolenia krzyżującego, przenieść średnią postaci z wartości 0 do wartości b, jeśli wybierze osobniki o wartości a. możemy pokazać, że b = h2a. aby wybrać cechę, warto zwiększyć współczynnik dziedziczności poprzez standaryzację środowiska (zmniejszając w ten sposób wariancję środowiskową) i umieszczenie się w warunkach środowiskowych sprzyjających pojawieniu się cechy., Genetyk Waddington wykazał, że w przyrodzie zmiana środowiska może w ten sposób ujawnić nowe cechy, które następnie zostaną wybrane podczas ewolucji.
Referencje i uwagi
obraz okładki. Muszle ślimaków Cepaea nemoralis & Cepaea hortensis. André Künzelmann, UFZ.,
wzór ten można zapisać w bardziej konwencjonalny sposób jako: \ pi = \ frac{n (n-1))} {2L}\sum _{i=1}^{N-1}\sum_{j=i+1}^{N}\delta ij
będąc próbką osób, u których zmierzono badaną cechę, wariancję można zdefiniować jako różnicę między średnią kwadratu wartości a kwadratem średniej wartości. Ta miara, która jest zawsze pozytywna, wskazuje na rozproszenie jednostek.
jest to dziedziczność w ścisłym tego słowa znaczeniu. Dziedziczność w szerokim znaczeniu to stosunek sumy wariancji genetycznych do wariancji całkowitej., Wzór ten daje dziedziczność z obojgiem rodziców („Przeciętny rodzic”); oszacowanie z tylko jednym rodzicem dałoby h2 / 2.
Encyclopedia of the Environment by the Association des Encyclopédies de l 'environnement et de l' énergie (www.a3e.fr), umownie związany z Uniwersytetem w Grenoble Alpes i Grenoble INP i sponsorowany przez Francuską Akademię Nauk.
przytoczyć ten artykuł: VEUILLE Michel (2021), polimorfizm genetyczny i zmienność, encyklopedia środowiska, url: https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
artykuły w Encyklopedii środowiska są udostępniane na warunkach licencji Creative Commons BY-NC-SA, która zezwala na powielanie pod warunkiem: powoływania się na źródło, Nie wykorzystywania ich do celów komercyjnych, udostępniania identycznych warunków początkowych, powielania przy każdym ponownym użyciu lub dystrybucji wzmianki o tej licencji Creative Commons BY-NC-SA.
Dodaj komentarz