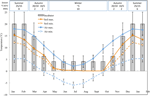
Figure 1. Dados mensais sobre a temperatura máxima e mínima do ar e do solo nos Alpes Australianos (média ± STDEV). As temperaturas do ar são médias da base de dados do BOM, as temperaturas do solo foram medidas com iButtons (ver métodos). As barras indicam temperaturas experimentais da incubadora (média ± STDEV)., A hora equivalente do ano( estação), o regime de temperatura da incubadora (°c, 12/12 h, luz/escuridão) e a duração do tratamento (semanas), conforme indicado no painel superior.
Com vista para descobrir as condições de temperatura próximos daqueles que sementes de experiência pós-dispersão, diária máxima e mínima, temperatura do solo os dados foram coletados no local de estudo (Marinheiro da Cabana; 36.27°S e 148.17°E, 2030 m um.s.l.) usando o ibutton registradores de dados (n = 10, Incorporado Sistemas de Dados, EUA) colocou de 4 cm abaixo da superfície do solo na base da vegetação a partir de 17 de janeiro a 17 de dezembro de 2012., Colocámos madeireiros a 4 cm para evitar perturbações da superfície, mas continuamos a ser representativos das condições a que as sementes são expostas no solo; esta profundidade é intermédia ao que foi utilizado nos anteriores exercícios de monitorização da temperatura do solo alpino (Scherrer e Körner, 2011; Pauli et al., 2015). Como se suspeita, a temperatura média do solo não desceu abaixo do congelamento durante o inverno (Figura 1). No verão, a temperatura do solo sob vegetação é conhecida por acompanhar a temperatura ambiente, enquanto as temperaturas do solo nu excederão o ambiente em até 15 ° C (Serviço de conservação do solo não publicado, 1960-1970).,as sementes maduras de 54 espécies de 16 famílias e 37 géneros foram recolhidas entre janeiro e abril de 2009, 2010 e 2011 (Ver quadro 1 Para nomes completos e autoridades). Os vales foram alojados no Herbário Nacional Australiano, Canberra). No total, a espécie representava mais de um quarto da flora angiosperma Australiana encontrada em regiões alpinas (Costin et al., 2000, Ver Tabela 1), embora muitas destas espécies se estendam até abaixo da linha das árvores também., A viabilidade de todas as coleções foi estimada antes da sementeira em condições experimentais de germinação usando a técnica de coloração do cloreto de tetrazólio (TZ) (International Seed Testing Association, 2003). Para mais pormenores sobre a recolha e o processamento, ver o Apêndice A em material suplementar.
a Tabela 1. Espécies de estudo, família e autoridade, forma de crescimento e detalhes das coleções, percentagem de viabilidade das sementes (TZ-estimada, ± E.), percentagem final de germinação alcançada (± S. E.,), diferença estatística entre viabilidade e germinação (G < V, testada apenas nos casos em que a germinação era inferior à viabilidade).a fenologia da germinação de sementes foi investigada imitando regimes de temperatura que as sementes alpinas experimentam in situ, pós-dispersão, numa progressão artificialmente reduzida das estações. As sementes foram imbibidas durante todo o experimento, portanto os resultados indicam potencial para germinação quando a água não é limitante., Os dados sobre a temperatura do ar e do solo (Figura 1) foram utilizados para orientar e validar os regimes de temperatura da incubadora, embora os logísticos contrastes de trabalhar com incubadoras impedissem a incorporação das flutuações inerentes às condições naturais. Os testes de germinação utilizaram oito réplicas de 25 sementes por colheita, semeadas em placas de Petri de plástico de 9 cm de diâmetro, contendo 1% de ágar-água simples. As placas de Petri foram seladas usando Parafilme para evitar a dessecação de ágar, antes de serem colocadas em incubadoras de germinação (Thermoline Scientific, Melbourne, NSW, Austrália)., Metade dos pratos foram embrulhados em folha de alumínio para excluir a luz e fornecer uma indicação de requisitos de luz para a germinação. Cada uma das quatro réplicas por tratamento de luz por colecção foi colocada numa prateleira diferente dentro da mesma incubadora e a localização dentro dessa prateleira foi re-arranjada semanalmente. Todas as réplicas de cada coleção foram movidas através dos seguintes termoperíodo simulado: 8 semanas a 25/15°C (Temperatura de dia/noite) → 2 semanas a 20/10°c → 2 semanas a 10/5 ° C → 10 semanas a constante 4-5 ° C → 2 semanas a 10/5 ° c → 2 semanas a 20/10°C → 6 semanas a 25/15 ° C., Assim, um ciclo de toda a experiência imitou as temperaturas que lembram o verão, o início do outono, o final do outono, o inverno, o início da primavera, o final da primavera e o verão, e durou 32 semanas no total. O ano abreviado foi considerado razoável porque as adesões geralmente tinham cessado a germinação em um determinado tratamento antes de ser deslocado para a próxima “temporada”., Uma 12/12 h luz/escuro fotoperíodo foi fornecida por lâmpadas fluorescentes (> 90 µmol m−2 s−1 no centro de cada prateleira), e na primavera, verão e outono regimes de temperatura de luz, coincidiu com a temperatura morna período do dia. A germinação, definida como a emergência de radicle em mais de 1 mm, foi marcada a cada 7 dias, e sementes germinadas e mudas foram posteriormente removidas dos pratos. A germinação das placas envoltas em folhas foi avaliada como acima, mas sob uma luz verde muito baixa.,
Após um ciclo experimental (32 semanas), a maioria das colecções foram terminadas (ver Informação suplementar para excepções), tendo demonstrado germinação zero durante pelo menos 2 semanas neste momento. Após o término, todas as sementes intactas remanescentes foram dissecadas com um bisturi sob um microscópio. As sementes com um endosperma firme e fresco e o embrião foram considerados viáveis e as sementes vazias de um embrião foram deduzidas do total ao calcular a percentagem de germinação para cada colheita., Calculámos um índice dos Requisitos de germinação da luz para cada espécie como uma fracção da germinação no tratamento escuro no momento em que os percentis cumulativos de germinação de 50 e 95% foram atingidos no tratamento Alternante luz/escuridão (ver abaixo).compilámos uma base de dados de características funcionais e medidas geográficas das espécies a partir de dados publicados para avaliar correlações com a estratégia de germinação., A base de dados incluía os seguintes traços contínuos: massa média das sementes, necessidade de luz para germinação (ver acima), área específica da folha (área fresca/massa seca), área individual da folha, peso individual seco da folha e altura na maturidade. As elevações mínimas, médias e máximas de coleta para a espécie foram determinadas a partir de todos os registros herbários no Herbário Virtual australiano, e a elevação em que as coleções específicas de sementes utilizadas no estudo foram feitas também foi incluída como traços contínuos. A forma de crescimento foi marcada como uma das três categorias: graminóide, herb, ou arbusto., Nós também incluímos uma característica binária: se as sementes eram endospérmicas ou não.
os dados do traço foram coletados de uma variedade de fontes, incluindo o Herbarium Virtual Australiano (http://avh.chah.org.au/), O Australian National Herbarium specimen information register (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Alpine Flora (Costin et al., 2000), The New South Wales Flora (Harden, 1990), and the comparative internal morphology of seeds (Martin, 1946). A massa de sementes e a área específica da folha foram obtidas a partir de coleções de campo individuais utilizadas nos ensaios de germinação., A massa de sementes foi obtida pesando quatro lotes de 10 sementes. O SLA foi determinado com base em cinco folhas totalmente expandidas de indivíduos separados por espécie. Estes foram coletados em campo, digitalizada em um scanner de mesa, seca a 60°C até peso constante e ponderada para o mais próximo 0.000 g. Onde insuficiente semente estava disponível, ou específicos da área da folha não estava disponível, os dados foram obtidos a partir de outras ANBG coleções da mesma espécie ou de outras fontes de dados (S. Venn, dados não publicados)., Se uma semente era endospermica ou não foi determinada a partir de desenhos da semente nos registros do banco de sementes da ANGB em consulta com o pessoal do banco de sementes da ANBG. A base de dados completa e as fontes estão disponíveis A pedido dos autores.
A análise de dados
os padrões de germinação foram avaliados visualmente e usando uma análise de cluster em PC-ORD para Windows, versão 6 (MJM Software Design, Gleneden Beach, ou, EUA). The analysis used a Euclidean distance matrix and a nearest neighbor joining algorithm., Os pontos de dados para a análise de clusters foram o tempo (semanas) para percentis específicos de germinação cumulativa (t20, t40, t50, t60, t80 e t95%) para cada coleção (média em 4 pratos). Assim, uma coleção que germinou no início da experiência teria curtos períodos de percentil até t80%, por exemplo, enquanto uma coleção que adiou a germinação até que as temperaturas aumentassem após o inverno simulado teria longos tempos para esses percentis., Padrões de germinação mais complexos, tais como a germinação lenta, constante ou explosões de germinação, também poderiam ser detectados, e seriam indicados por tempos uniformemente espaçados ou vezes agrupados no início e no final do experimento, respectivamente. Este método foi usado como uma forma de impor uma avaliação objetiva do padrão de germinação, independente da temperatura per se.,foi realizada uma análise de Sentido Único da variância (ANOVA) para as placas replicadas de cada coleção, a fim de avaliar se havia uma diferença significativa entre a viabilidade percentual (determinada pelo teste TZ) e a germinação percentual final. Foram utilizados outros modelos lineares gerais para avaliar se os agregados resultantes da análise dos agregados diferiam em termos de Viabilidade ou de percentagem final de germinação.associações entre traços ecológicos e clusters de estratégia de germinação foram testadas de duas maneiras., Em primeiro lugar, testamos diferenças médias entre clusters usando ANOVA com cluster como um efeito fixo e correlatas ecológicas como as variáveis de resposta, ou usando G-testes replicados para endospermia e forma de crescimento. Em segundo lugar, porque a germinação e outros traços funcionais são altamente susceptíveis de ser influenciados pela história evolutiva, nós avaliamos essas correlações usando regressões filogenéticas. A phylogeny was constructed for the study species using Phylocom Phylomatic and the bladj packing in R (Webb et al., 2008; R Core Team, 2013)., Nós avaliamos a presença da estrutura filogenética em variáveis usando o phylosig ou phylo.função d (variáveis discretas ou binárias) do pacote Fitools em R (Revell, 2012). As regressões filogenéticas foram conduzidas usando a função pgls em Caper (Orme et al., 2013). As análises foram realizadas de forma estruturada, reflectindo a agregação revelada no PC-Ord (ver infra)., Inicialmente incluímos quatro estratégias germinativas (adiadas, imediatas e escalonadas dentro de cada uma das anteriores), depois três (agrupando todas as espécies escalonadas e comparando com adiadas e intermediárias) e finalmente avaliamos as diferenças entre as duas estratégias mais amplas, adiadas e intermediárias.a viabilidade das sementes em comparação com a germinação Final em geral, a viabilidade das sementes e a percentagem final de germinação foram elevadas. A viabilidade média da TZ estimada em todas as colecções foi de 80 ± 1.,7%, com mais de dois terços das coleções exibindo mais de 75% de viabilidade (Quadro 1). Não houve diferença na qualidade das coleções através de 3 anos (média de viabilidade em 2009, 2010 e 2011 foi de 79 ± 2.8, 80 ± 2,7 e 81 ± 3.7%, respectivamente), indicando que a banca de sementes coletadas em 2009 não diminuir a sua viabilidade.germinação que excedeu ou não foi significativamente inferior à viabilidade estimada da TZ foi atingida para 45 das 54 espécies (ANOVA de Sentido Único: P>0,05, Quadro 1)., Das nove espécies restantes, a germinação final de quatro espécies foi muito baixa (< 5%), apesar da boa viabilidade (ca. 55-94%), pelo que não conseguimos tirar quaisquer conclusões sobre as estratégias de germinação destas espécies. A incapacidade de germinar poderia refletir que nossos tratamentos não aliviaram a dormência ou que as sementes não estavam totalmente maduras na coleta. Testes de corte no final da experiência indicaram que, em todos os casos, a maioria das sementes não germinadas continha um embrião saudável e imbuído., Curiosamente, 34 espécies atingiram uma germinação superior à viabilidade estimada da TZ (Quadro 1), sugerindo que o teste da TZ subestimou frequentemente a viabilidade das sementes.
aglomerados de germinação
a análise de aglomerados revelou dois aglomerados principais (A E B), com subestrutura em cada (figura 2). Para a maioria das espécies (n = 21 espécies de 10 famílias), a germinação foi adiada para um período de temperaturas de Inverno (5°C dia/noite) ou até que as temperaturas foram aumentadas para 10/5°C após o inverno (A1 e A1:inverno, Figuras 2, 3A)., Alternativamente, a germinação começou imediatamente a 25/15 ° c (n = 17 espécies, seis famílias, vistas no aglomerado B1, Figuras 2, 3B). Para a maioria destas espécies, a taxa de germinação foi rápida; 50% da germinação total (t50) ocorreu em menos de 3 semanas (figura suplementar 1).
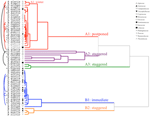
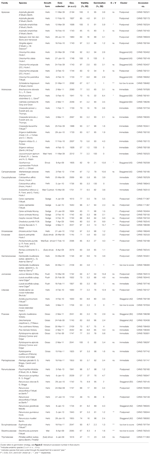
Figura 2. Análise de clusters dendrogram baseado no tempo para o percentis cumulativo de germinação (20, 40, 50, 60, 80, 95%). Espécies indicadas por género inicial e primeira parte do nome da espécie. Asterisks marcam coleções que foram banked antes do teste., Os arcos finos conectam coleções duplicadas para comparar o efeito do banco na germinação. Símbolos denotam famílias diferentes (ver legenda). Coleções mostradas em texto cinza são aquelas que foram expostas a dois ciclos do experimento.
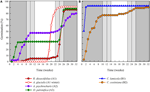
a Figura 3. Germinação percentual cumulativa de uma espécie representativa de cada agregado germinativo. (A) cluster a, species that adied germination, (B) cluster B, species that germinated immediately., Note que os padrões de germinação escalonados são aninhados dentro dos aglomerados acima. Os regimes de temperatura da incubadora são representados por sombreamento (ver Figura 1).
dentro de cada uma destas classes amplas, no entanto, houve sub-clusters claros que refletiam a germinação que foi escalonada ao longo do tempo, ou seja, uma proporção do lote de sementes germinou antes da exposição às temperaturas do inverno, enquanto o restante germinou após o inverno. A germinação escalonada ocorreu em 13 espécies de seis famílias e ocorreu principalmente quando as temperaturas diurnas excederam 10 ° C (figuras 3A,B)., Estes germinadores escalonados foram divididos em três grupos na análise de clusters. O Cluster A2 com sete coleções mostrou germinação substancial tanto antes quanto depois do período de temperatura fria. Quando a germinação cessou antes de As temperaturas terem sido alteradas, concluímos que a falta de outra germinação antes do inverno não foi motivada pela temperatura (A3). Em contraste, a germinação de grande parte do Grupo A2 parecia ser interrompida apenas quando as temperaturas foram reduzidas para 10/5 ° C., O aglomerado B2 continha mais quatro espécies que exibiam uma germinação substancial no início do experimento e passou a alcançar uma proporção relativamente pequena de sua germinação total após as temperaturas serem retornadas para acima de 5°C (figura 3B).em comparação com a variação relativamente menor entre as espécies dentro dos aglomerados A1 E B1, os aglomerados escalonados também mostram uma diferenciação muito maior entre as espécies. As curvas individuais para todas as espécies são apresentadas nas figuras suplementares 1i-VII. para mais pormenores sobre a estratégia de germinação, ver Apêndice B em material suplementar.,
Ecológica Correlatos de Germinação Estratégia
De todos os ecológica correlatos que nós considerado, apenas aqueles diretamente associados a sementes de traços foram significativamente correlacionados com a germinação de estratégia, independentemente de se avaliou-se a correlação ao nível de dois grandes clusters (adiado vs. imediata de germinação), ou ao nível de três ou quatro clusters, ou se nós representaram filogenética estrutura ou não (Tabela 2)., As espécies com germinação adiada tinham sementes mais pesadas e eram mais propensas a ser endospérmicas em comparação com as espécies que germinavam imediatamente (Figura 4). Espécies com germinação cambaleante eram intermediárias na massa de sementes e endosperma. No entanto, nem todas as características das sementes estavam correlacionadas com a estratégia de germinação: não havia correlação entre os requisitos de luz para a germinação das sementes e a estratégia. Contrariamente às nossas expectativas, as espécies com uma área de folha específica mais elevada (potencialmente indicativa de taxas de crescimento mais elevadas) não apresentam maior probabilidade de germinação imediata., Da mesma forma, espécies com elevação média mais elevada ou intervalos de elevação menores não eram mais propensos a exibir germinação adiada.

Tabela 2. Incidence of phylogenetic structure in, and correlates of, germination strategy.
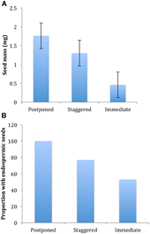
Figura 4., Diferenças na (a) massa média de sementes e (B) proporção de espécies com sementes endospérmicas entre espécies com estratégias adiadas (cluster A1), escalonadas (combinação dos clusters A2, A3 e B2) e germinação imediata (cluster B1). As diferenças entre as estratégias são significativas em P ≤ 0,05, ver Quadro 2.
discussão
germinação em habitats alpinos foi historicamente considerada como ambientalmente controlada, com sementes de Inverno isolando a neve contra potenciais sinais de germinação e tornando a dormência desnecessária (Billings and Mooney, 1968)., No entanto, os nossos resultados apoiam provas mais recentes de que os mecanismos de dormência são comuns entre as espécies alpinas, tanto na Austrália (Hoyle et al., 2013; Sommerville et al., 2013) e em outras partes do mundo (Cavieres e Arroyo, 2000; Schutz, 2002; Shimono e Kudo, 2005; Mondoni et al., 2009; Schwienbacher et al., 2011)., A dormência parece desempenhar um papel significativo no controle do tempo in situ de germinação de muitas espécies alpinas australianas, atuando para atrasar, adiar ou retardar a taxa de germinação e, assim, potencialmente conferindo estratégias de regeneração avessa ao risco em um ambiente duro e variável. Talvez o mais marcante de nossos resultados de germinação foi a proporção de espécies para as quais a estratégia de germinação variou dentro de uma coleção de sementes para uma determinada espécie, sugerindo que as características dormentes e não dormentes foram exibidas dentro da mesma coleção de sementes destas espécies., Notavelmente, as estratégias de germinação eram geralmente altamente conservadas dentro das famílias e também correlacionadas com elementos da anatomia das sementes: massa e endospermia. As estratégias de germinação foram, no entanto, independentes de outros traços funcionais ecologicamente significativos da planta madura.as sementes de quase metade das espécies estudadas pareceram incapazes de germinar até serem expostas a um período fresco e húmido (constante 5°C), sugerindo que a estratificação fria aliviou um mecanismo de dormência fisiológica., Adiar a germinação até a primavera seguinte pode permitir que as mudas evitem estabelecer-se sobre ou antes do inverno rigoroso, ao mesmo tempo que otimiza o curto período de crescimento que se aproxima. Em contraste, a germinação oportunista exibida pela espécie que germinou imediatamente pode proporcionar uma vantagem seletiva quando o risco de mortalidade de plântulas de inverno é baixo, permitindo que as plantas florem mais cedo na primavera seguinte ou em maior tamanho (Donohue, 2002).,até à data, tem sido publicada pouca evidência de variabilidade intra-específica nos mecanismos de dormência dentro das coleções de sementes alpinas, tal como explicaria as estratégias escalonadas de germinação observadas aqui. A germinação escalonada ao longo do tempo pode também ser explicada por diferentes níveis de maturidade das sementes entre os indivíduos da população no momento da colheita, e/ou pode refletir uma estratégia dimórfica dentro de uma única planta associada à posição na planta ou momento do desenvolvimento.,a bet-hedging evolucionária é frequentemente evocada para explicar a estratégia diversificada em características de germinação de sementes (Simons, 2011), como a demonstrada pelas coleções com germinação escalonada. Se, no entanto, uma estratégia diversificada resulta em um aumento da aptidão média e uma redução na variância (por exemplo, reduzindo a concorrência entre irmãos), então não constitui bet-hedging. Da mesma forma, se a aparente diversificação reflete diferenças entre os indivíduos, não dentro, então não é bet-hedging (Starrfelt e Kokko, 2012)., As investigações sobre os padrões de dispersão das plântulas alpinas e a tolerância à geada e ao degelo, associadas a estudos sobre as determinantes da variação da estratégia de germinação dentro das espécies, podem ajudar a explicar a fenologia da germinação das espécies que exibem uma estratégia escalonada e podem indicar se são susceptíveis de representar estratégias de Bet-hedging.,
correlatos ecológicos da Estratégia de germinação
nosso exame da relação entre a estratégia de germinação e outros traços ecologicamente importantes demonstrou que os traços de germinação estavam correlacionados com outros traços de semente, mas não com traços vegetativos. Sommerville et al. (2013) found that Australian alpine herbs with non-endospermic seeds were more likely to be non-dormant at dispersal. Nosso trabalho apoia essa descoberta e indica ainda que as sementes menores são mais propensas a estar entre os germinadores imediatos. Embora Sommerville et al., (2013) não postulou por que endosperma pode estar associado com dormência, sugerimos que a colocação de reservas em cotilédones pode melhorar a taxa de crescimento precoce e Estabelecimento Em espécies germinativas imediatas. Os nossos resultados não indicaram qualquer correlação entre a necessidade de luz e dormência, embora os requisitos de luz variassem entre as espécies. Há uma variedade de outros traços de sementes que protegem o risco de extinção em ambientes variáveis, Por exemplo, mecanismos de dispersão de sementes e longevidade de sementes no banco de sementes do solo (ver Tielborger et al., 2012 para uma revisão)., Nesta fase, não podemos dizer se essas características mostrariam ou não correlações com a estratégia de germinação.mais tarde perguntamos se a estratégia de germinação estaria correlacionada com traços não-reprodutivos ecologicamente importantes, como a área específica da folha, ou a altura da planta (Westoby, 1998; Wright et al., 2004). Em particular, postulamos que as espécies germinativas imediatas podem apresentar valores de traços indicativos de taxas de crescimento mais rápidas (usando SLA como um proxy) que podem ser necessárias para o estabelecimento no curto período entre a dispersão e o início do inverno., No entanto, não encontramos tais correlações. Se os traços fisiológicos e de crescimento das mudas nos estágios iniciais do Estabelecimento podem mostrar padrões diferentes do que os traços vegetativos adultos é uma questão digna de consideração, mas além do escopo do presente trabalho.
Finalmente, examinamos se as espécies de altitudes mais elevadas, onde épocas de cultivo são geralmente mais curtas podem ser mais propensos a apresentar dormência, mas não houve associação entre a elevação da faixa, coleta de elevações, ou a altitude média da coleção para a espécie como um todo e de germinação de estratégia., Juntos, estes resultados indicam que, embora a evolução da estratégia de germinação esteja intimamente ligada a traços de sementes ecologicamente importantes, ela compreende um eixo em grande parte independente às características vegetativas ou distributivas.os ecossistemas alpinos australianos, como as áreas alpinas em todo o mundo, estão sob ameaça de alterações climáticas combinadas com mudanças na frequência de incêndios, padrões de uso da terra, influxo de espécies invasoras e os impactos do aumento da visitação humana., O aquecimento associado às alterações climáticas está a ocorrer mais rapidamente acima da linha das árvores do que em elevações mais baixas, e prevê-se que as zonas alpinas continuem a sofrer um aquecimento superior à média no futuro (Kullman, 2004). A capacidade de Regeneração contínua através de sementes em novas condições é susceptível de desempenhar um papel significativo na resposta das plantas alpinas às alterações climáticas e às alterações associadas. Além disso, é provável que os regimes de perturbação alterados exijam um papel acrescido de restauração e Reabilitação na região., Os insights fornecidos aqui são, portanto, cruciais para a conservação e gestão dos sistemas alpinos em processo de mudança; eles podem ajudar a informar nossas previsões de como as espécies de estudo podem ser afetadas, formar a base de planos de propagação de sementes para estas espécies, e ser usados para orientar futuras investigações sobre estratégias de germinação de sementes alpinas com resultados identificáveis de conservação e gestão.,
As alterações de temperatura são susceptíveis de afectar o estado de dormência das sementes tanto pré como pós-dispersão, alterando e / ou perturbando as estratégias de germinação e resultando em mudanças na fenologia actual da germinação (Hoyle et al., 2008b). Para as plantas alpinas, o aquecimento climático pode levar a uma mudança da primavera para o outono, impulsionado principalmente por alterações no estado de dormência das sementes e resultando em importantes implicações para as espécies atualmente adaptadas ao surgimento na primavera (Mondoni et al., 2012)., Mais pesquisas sobre como mudanças nas temperaturas que as sementes experimentam pós-dispersão, redução da duração da cobertura de neve, e aumento da frequência de eventos de temperatura extrema vai afetar a germinação são necessárias, a fim de prever como as espécies com dormência vai responder à mudança climática. Para as espécies alpinas que germinam no outono, antes da geada e da queda de neve, os nossos resultados levantam a questão de como as mudas destas espécies irão lidar com a redução prevista da precipitação e com o aumento da frequência dos eventos de geada sazonais.,finalmente, os nossos resultados levantam questões importantes sobre a variação dentro das espécies, particularmente em associação com a previsão do impacto das alterações climáticas ou a selecção das sementes a utilizar para a restauração. As estratégias de germinação podem variar dentro das espécies dependendo da população, tempo de floração, massa de sementes, investimento parental, clima, elevação e/ou ambiente materno e esta variação pode refletir fatores genéticos, epigenéticos e/ou ambientais (Cochrane et al., 2015)., Increasing elevation accounts for variation in germinability and / or dormancy status of several alpine species (Billings and Mooney, 1968; Dorne, 1981; Cavieres and Arroyo, 2000) but see (Hoyle et al., 2014). O estado de dormência reduzido pode resultar do término antecipado do desenvolvimento de sementes resultante de tensões ambientais (Steadman et al., 2006; Hoyle et al., 2008b)., O exame da variação das estratégias de germinação dentro das espécies pode revelar o potencial das espécies para lidar com e adaptar-se às mudanças climáticas, desempenhando assim um papel importante na determinação da sobrevivência futura e da distribuição das espécies e proporcionando uma visão importante para a gestão e conservação.
conclusões
nossa abordagem revelou a ocorrência generalizada de dormência na flora Alpina australiana e demonstra que as estratégias de germinação variam dentro de uma coleção de sementes, bem como entre coleções., Além disso, temos mostrado que enquanto a estratégia de dormência é filogeneticamente conservada e correlacionada com a massa de sementes e endosperma, ela é em grande parte independente de traços vegetativos e características de gama. Embora não se possa extrapolar directamente dos nossos resultados a forma como o clima e as alterações associadas irão afectar a flora Alpina, estes resultados podem constituir a base para a concepção de planos de propagação e para novas experiências sobre o impacto das alterações dos regimes climáticos., Em particular, defendemos uma investigação mais aprofundada sobre a compreensão das causas da variação dentro das espécies na estratégia de germinação e o papel que esta pode desempenhar na capacidade das espécies para resistir às alterações climáticas previstas.
Data Accessibility
Full germination data and trait database will be lowed on DRYAD and with the Atlas of Living Australia.
Declaração de conflito de interesses
os autores declaram que a investigação foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.,reconhecemos o financiamento do Australian Research Council (LP0991593) e do Australian National Botanic Garden e seus amigos. Agradecemos a J. McAuliffe, S. Fethers, e S. Lawatsch do ANBG. Agradecemos também a M. Holloway-Phillips, M. Cardillo, E S. Cunningham pela entrada em versões anteriores do manuscrito e por aconselhamento analítico e S. Venn pelo acesso a dados de traços.
material suplementar
Foley, M. E. (1992). Efeito dos açúcares solúveis e do ácido giberélico na quebra da dormência de excitados embriões de aveia selvagem (Avena fatua)., Weed Sci. 40, 208–214.
Google Scholar
Harden, G. J. (1990). Flora of New South Wales. Sydney, NSW: Royal Botanic Gardens.
Google Scholar
International Seed Testing Association. (2003). Folhas de trabalho sobre os testes de Tetrazólio. ISTA.
Kullman, L. (2004). A face em mudança do mundo alpino. Glob. Muda O Noticiário. 57, 12–14.
Google Scholar
Deixe uma resposta