
polimorfismo genético é a existência de Estados Alternativos de DNA, determinando a variação dos níveis mais elevados de integração do organismo. Existem diferentes tipos de modificações do genoma (mutações). Os mais estudados são substituições nucleotídicas em regiões de codificação e regulação.
Definição
o polimorfismo Genético é a existência, em uma população, de várias alternativas de DNA, ou alelos, em uma posição definida no genoma, ou locusPosition do gene no cromossomo., Em genética populacional, um conjunto de genes homólogos (classe de homologia). Dois genes são homólogos se coincidirem com a meiose. Plural: loci.. Esta definição tem vários aspectos:
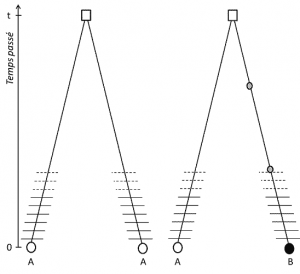
(1) em primeiro lugar, o traço deve ser carregado pelos cromossomos e ser transmissível.
(2) em seguida, os alelos devem ser homólogos de dois cromossomos ou dois genes são ditos homólogos se eles coincidem e mutuamente excluem um ao outro da meiose. pela sua posição no genoma, que exclui certos tipos de variação que serão discutidos mais tarde.,
(3), Mas desde que o traço é transmissível, posição homologia também implica que os alelos são homólogos por descendência; se eles são diferentes, portanto, é que uma mutação (pelo menos) ocorreu em uma das linhas que os ligam ao seu último ancestral comum (Figura 1).
(4) Finally, genetic polymorphism can be defined at the scale of the smallest unit composing DNA: the nucleotide site., Como resultado, cada variante do nucleótido pode determinar o polimorfismo dos níveis mais elevados da estrutura biológica – gene, proteína e fenótipo do indivíduo – que se tornam escalas nas quais o polimorfismo genético pode ser descrito (Figura 2).
tome o exemplo do sistema sanguíneo ABO, que corresponde à parte antigénica do antigénio reconhecida por um anticorpo ou receptor linfocitário. Também conhecido como um epítopo ou um determinante antigênico. O mesmo antígeno pode ter vários epítopos (idênticos ou diferentes) e assim induzir uma resposta imunitária variada., envolvido na compatibilidade durante as transfusões sanguíneas. Este é um caso de polimorfismo genético presente em populações humanas, envolvendo a proteína ABO glicosil transferaseenzima permitindo a transferência de resíduos contendo açúcares para proteínas. No caso do sistema ABO, a glicosil transferase a e B, respectivamente, induzem um indivíduo a pertencer ao grupo A ou B. Quando ambas as glicosiltransferases estão presentes, o indivíduo é do Grupo AB. que tem três alelos, A, B E O. Um indivíduo pode ter (AA), (AO), (BB), (BO), (OO) ou (AB) genótipos., Seu fenótipo será então ,,,;,; ou, onde vemos que a e B “dominam” O (isto é, sua expressão mascara a de O), e que eles são co-dominantes entre si (o heterozigótico entre A e B tem uma expressão reconhecível,).
um polimorfismo pode ser descrito primeiramente na escala da sequência de DNA do locus que codifica uma proteína. Alguns polimorfismos são “sinônimos”, ou seja, eles não mudam a sequência de aminoácidos da proteína; eles são muitas vezes os polimorfismos mais numerosos. Os outros modificam aminoácidos e são chamados de polimorfismos de “substituição”., Polimorfismos de substituição da ABO glicosil transferase são de dois tipos: polimorfismos que mudam o aminoácido, mas não a unidade antigênica (eles não interferem com o fenótipo ABO dos indivíduos); e outros polimorfismos de substituição, que determinam o abo fenotipeset de características observáveis de um indivíduo..
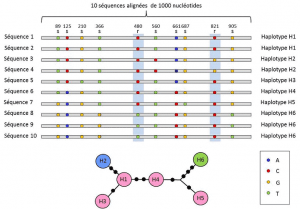
entre os níveis do nucleótido e do alelo, as análises genéticas têm em conta um nível intermédio de descrição: A disposição linear de sítios variáveis no local. Estes arranjos, chamados de haplotipesgrupo de alelos de diferentes loci localizados no mesmo cromossomo e geralmente transmitidos juntos. Haplotype é um genótipo haplóide formado pela contração da frase inglesa “genótipo haplóide”., Todos os genes localizados no mesmo cromossoma e cujos alelos se segregam juntos durante a meiose constituem um haplótipo. Estes genes são chamados de “geneticamente ligados”, são úteis na genética evolutiva porque permitem encontrar as ligações genealógicas entre os alelos (Figura 2).polimorfismos de nucleótidos também existem em regiões intergênicas (fora das regiões de codificação). Alguns afetam as áreas onde a expressão genética é regulada e, portanto, têm expressão fenotípica. Outros não têm nenhum efeito conhecido e são referidos como “silenciosos”.,
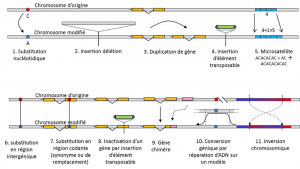
além de substituições de nucleotídeos, polimorfismo para um tipo específico de mutação, microssatélites, refere-se a variações no comprimento de uma, muitas vezes não-codificantes do DNA do fragmento (Figura 3), devido a uma alteração no número de repetições em um curto espaço de repetidas seqüência de nucleotídeos, e.g. CACACACA, ou TGTGTG.
nem todas as variações genéticas são abrangidas pela definição de polimorfismo genético, uma vez que requer a homologia de posição de alternativas (Figura 3)., Assim, há sequências repetidas obtidas pela duplicação de um gene, para o qual duas repetições não são homólogas por posição. Muitas vezes, também não podemos falar de homologia para elementos transponíveis, porque eles geralmente mudam de posição durante as gerações celulares, podem multiplicar e invadir o genoma. A homologia de dois elementos transponíveis não pode então ser definida. Por outro lado, duas sequências de codificação do mesmo locus, uma das quais está intacta, e a outra inactivada pela inserção de um elemento transposível, correspondem bem a uma situação de homologia., Um dia, vamos provavelmente considerar falar sobre polimorfismo para as modificações dos chamados cromossomas “epigenéticos”, que são por vezes transmitidos ao longo de várias gerações celulares, somaticqualifica células não-reprodutivas, ou soma. Uma mutação somática, que afeta um gene de uma célula somática, desaparece com o portador individual. ou germinalqualifica gâmetas. Uma mutação germinal pode ser transmitida à sua descendência., tais como metilação. A sua importância na evolução das populações permanece por avaliar., A limitação da noção de polimorfismo genético a certas categorias de variações – essencialmente substituições nucleotídicas – vem da grande utilidade deste último na exploração da história das populações naturais. Eles se prestam à modelagem matemática da evolução.
A Palavra polimorfismo significa “várias formas”. É contrário ao monomorfismo, o que indica a ausência de variação. No vocabulário da biologia, esta oposição tem sido usada há muito tempo, antes da genética, para sinalizar a coexistência de vários tipos de indivíduos na mesma espécie, e.g., polimorfismo de castas (rainha e trabalhadores) em insetos sociais; polimorfismo sazonal (mudanças de pele) em alguns mamíferos árcticos. Estes casos não se enquadram no polimorfismo genético, um conceito mais recente com um significado mais estreito e mais preciso. Variações contínuas (por exemplo, variações de tamanho) também não se enquadram na categoria de polimorfismo, uma vez que não apresentam alternativas distintas. No entanto, locus que afetam o tamanho estão dentro desta definição., Discutiremos abaixo as variações genéticas mais estudadas porque elas são as mais importantes para a evolução, substituições nucleotídicas, e então discutiremos a variação dos fenótipos.
medição
não é suficiente dizer que um locus é mais ou menos variável sem dar uma estimativa quantitativa a esta sentença. Um pesquisador pode estudar a variação em diferentes escalas. Se ele só está interessado nos alelos de uma proteína, ele vai medir a diversidade alélica, simbolizada por “H”. Se ele está interessado na diversidade de DNA, ele vai medir a diversidade nucleotídica, simbolizada por “π”.,
O alelo de diversidade H é definida como a probabilidade de sortear dois diferentes alelos durante dois amostragem com replacementPerforming sucessivas desenhar com a entrega da p tokens em uma urna contém n tokens, significa dar um primeiro token, a leitura de seu valor, colocá-lo novamente na urna, tendo um segundo token, a leitura de seu valor, colocá-lo novamente na urna, etc., até ao sinal de pth. Isso significa escolher objetos p entre n Com repetição (você pode escolher o mesmo objeto várias vezes) e por ordem (a ordem em que você escolhe os objetos é importante)., O número de sorteios sucessivos com tokens entre n é: n × n × n × … × n = np.. Se chamarmos pi a frequência do alelo do posto i, mostramos que a probabilidade de escolher o dobro do mesmo alelo é f = Σ pi2. A diversidade alélica da amostra é, então, o seu complemento para 1, i.e.:
H = 1 – ∑ pi2 (1)
Isto aplica-se tanto a proteína de alelos e haplótipos. Isto é chamado de diversidade haplotípica. Também pode ser chamado de heterozigosidade, porque no caso de um locus diplóide, ele dá a frequência esperada de heterozigotos.,
a diversidade nucleótida π é o equivalente de H na escala de cada nucleótido. É calculada como a média do número de diferenças de nucleótidos entre sequências de uma amostra tomada duas por duas (δij), dividido pelo comprimento do fragmento de ADN no número de nucleótidos (L).
π = média (δij)/L (2) (ver referência )
Este valor difere entre as espécies. Nas partes codificantes do genoma humano, dois cromossomas escolhidos aleatoriamente da população diferem em média por um nucleótido por mil., Na mosca da fruta (Drosophila melanogaster) esta diferença é de cerca de uma em cada cem. A mosca é assim dez vezes mais variável do que o homem. Quando estes valores estão relacionados com o tamanho das regiões codificantes (cerca de 15.500 os genes de Drosophila, pelo menos de 22.000 em seres humanos), ou mesmo de genomas (140 milhões de pares de bases por genoma haplóide em as moscas, cerca de 3.200 milhões ou vinte vezes mais em humanos), o número de sítios polimórficos é astronômico e não há um nascer do sexualizada geração de qualquer espécie é, tem sido e vai ser geneticamente idêntico a outro passado, presente ou futuro., O poder da Informação do polimorfismo do ADN é imenso. Nos forenses, os detectives podem identificar qualquer suspeito de 16 microsatélites.
História
O termo variação está no título dos dois primeiros capítulos do livro de Darwin A Origem das Espécies (1859). Darwin introduziu esta noção nas ciências naturais numa época em que as leis da hereditariedade biológica permaneciam um mistério, e dedicou-lhe outro livro importante, a variação de animais e plantas (1868)., Convencido de que a evolução era sobre pequenas variações que faziam pouca diferença na adaptação dos organismos às suas condições de vida, ele atribuiu importância crucial a Pequenas variações quantitativas, o que levou seus sucessores (notavelmente Karl Pearson) a encontrar a biometria da medição da vida. Refere-se, num sentido muito amplo, ao estudo quantitativo dos seres vivos.. Mas a redescoberta das leis de Mendel em 1900 mudou o interesse para variações descontínuas.
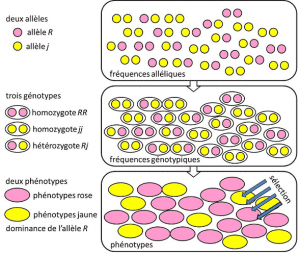
de 1908 a 1930, a genética populacional foi essencialmente uma disciplina teórica que procura reconciliar o darwinismo com a hereditariedade Mendeliana baseada na transmissão de um único gene sob um modo dominante, recessivo ou sex cromossomo X (ou Y). Refere-se a um traço genético com determinismo simples, por um casal ou um pequeno número de casais de genes., e onde as probabilidades desempenharam um papel importante. A genética é contra-intuitiva., Prevê que não há reprodução do progenitor em seu descendente, porque o genótipo deste último resulta da loteria da separação dos alelos antes da fusão dos dois meio genomas parentais. Foi percebido que estas são as freqüências alélicas em que o alelo de uma variante é encontrado em uma população. Expresso em percentagem ou proporção. A soma das frequências alélicas de todos os alelos de um gene em uma população é, portanto, por definição igual a 1. Em genética populacional, as frequências alélicas representam a diversidade genética a nível da população ou da espécie., que são transmitidos entre gerações, não genótipos ou fenótipos. Estas frequências são mais ou menos estáveis de uma geração para a outra e produzem, além do entrelaçamento múltiplo de filiações entre indivíduos, a mesma estrutura genotípica de freqüências genéticas da população. Determinada a partir das frequências alélicas (Figura 4). A distribuição de genótipos da população, associada à variância, é, portanto, o único fator previsível., Por volta de 1930, os três teóricos Ronald Fisher, JBS Haldane e Sewall Wright ajudaram a criar a noção da população Mendeliana cuja hereditariedade obedecia às leis de Mendel. . Neste contexto, a evolução associa necessariamente três níveis estruturais: o gene, o indivíduo e a população. Esta associação de três vias pode ser resumida pela seguinte fórmula: evolução da população é uma mudança na frequência de alelos (escala de genes, mas também escala de grupos), uma mudança que depende da classificação de fenótipos por seleção (escala individual)., Empírica genética de populações de investigação desenvolvido, mas foi limitado por um longo tempo por nossa ignorância de como os cromossomos de trabalho (estrutura do DNA foi entendido em 1953 e seu seqüenciamento começou em 1977), e tinha para voltar a cair no estudo dos poucos visível polimorfismos, tais como a coloração da joaninha do elytra ou o shell de caracóis (consulte o Foco, O grande, caracol debate). Houve muito debate entre os geneticistas sobre se as populações naturais eram geralmente polimórficas ou monomórficas, e se o polimorfismo era em si benéfico ou não., The study of macromolecular scale variation had to wait until 1966 (for proteins) and 1983 (for DNA) before answering these questions.variações quantitativas e hereditariedade quando olhamos à nossa volta, vemos diferenças físicas entre seres, alguns complexos como características faciais, outros fáceis de medir como peso ou altura. O bom senso sugere que são parcialmente transmissíveis, embora de uma forma difícil de especificar. Estes traços são muitas vezes poligênicos sob a dependência de muitos genes. Estamos a falar de hereditariedade poligénica. A Diabetes é uma doença poligénica., o que significa que são influenciados por muitos locus. Desenvolvimento da genética, e sua aplicação para a comparação das espécies (evo-devo”, para a “evolução do desenvolvimento”) desvendar a teia de interações complexas que fazem com que durante o desenvolvimento, as células do corpo, todos com a mesma bagagem genética, diferenciam-se pelo epigenesis para dar diferentes tecidos. Ainda não está claro como personagens complexos são construídos. Fenótipos mensuráveis podem ser estudados usando biometria, com base nas propriedades das distribuições estatísticas., Um valor com propriedades úteis é a variância em uma amostra de indivíduos em que um dado traço foi medido, a variância é a diferença entre a média do quadrado dos valores e o quadrado da média dos valores. Esta medida, que é sempre positiva, indica a dispersão dos indivíduos. . Quando várias causas independentes determinam a variação de uma linha, suas variâncias são aditivas e sua soma dá a variância da linha. Se não forem independentes, a soma das covariâncias é adicionada à soma das variâncias., A variância total Vt de uma característica fenotípica é tal que :
Vt = Vga+Vgd+Vgd+Vgi+Ve
Vga, genético aditivo variância é a soma dos desvios devido para cada locus, independentemente dos outros; Vgd, variância genética dominante, resulta da interação de dois alelos de um mesmo locus; Vgi, a interação variação entre locuses, resulta do fato de que o locuses o mesmo indivíduo interagir; Ve, a variância do meio ambiente, é assumido aqui independente da variância genética.,
a variância genética da dominância é comum a dois filhos dos mesmos pais, porque eles compartilham efeitos dominância quando em um determinado locus eles receberam os mesmos alelos de ambos os pais, o que acontece com eles em um em quatro locus. Eles não compartilham esses efeitos com seus pais, então eles são mais “similares” uns aos outros do que são com seus pais, apesar do fato de que eles compartilham tantos genes uns com os outros. Certamente, um pai pode, por acaso, ter recebido os mesmos dois alelos que um de seus filhos em um determinado locus., Esta possibilidade depende da frequência de alelos na população. Vemos que o grau de semelhança entre uma criança e seu pai para um determinado traço é uma propriedade pública.mais geralmente, todos os Componentes da variância genética (Vga+Vgd+Vgi) podem variar entre populações. Em linhas de reprodução, um caso comum é a consanguinidade, que esgota a variância genética: como resultado, um traço pode ter a mesma média em duas populações, mas tem uma variação genética diferente. A habilidade de um criador de selecionar um traço é, portanto, uma característica da linha, não do traço.,a relação entre pais e filhos é medida pela hereditariedade h2. Ele é definido como a proporção de aditivo genética de variância a variância total :
h2 = Vga/Vt

(2) correlação de 0, onde o tamanho dos filhos é independente do DOS Pais. Um pai médio valioso produziu um filho de valor indiferente, dentro da distribuição das crianças. As coordenadas dos pontos estão localizadas em um círculo; a expectativa de seu valor está localizada em uma linha paralela ao eixo dos pais (d)., (3) correlação mãe-filho intermediária entre 0 e 1: as coordenadas dos pontos estão localizadas em uma elipse. A expectativa de seu valor não está mais localizada no bissetor dos eixos de coordenadas (que também é o eixo principal da elipse de distribuição), mas na linha de regressão R. um pai médio de valor produziu um filho de Valor b (entre 0 e a) com uma certa variância. Note-se que o caso (3) é intermediário entre os casos (1) e (2)
é mostrado sob vários pressupostos na Figura 5., Hereditariedade é a capacidade de determinar se um recurso pode ser selecionado. A figura 5-3 mostra que um criador será capaz de, dentro de um cruzamento de geração, para mover a média de um personagem a partir do valor 0 para o valor de b se ele seleciona os indivíduos com valor um. Podemos mostrar que b = h2a. Para selecionar um traço, ele é útil para aumentar a herdabilidade relação com a padronização do ambiente (reduzindo assim ambiental variância), e colocando-se nas condições ambientais que favorecem o aparecimento de traços., O geneticista Waddington mostrou que, na natureza, uma mudança de ambiente pode, assim, revelar novas características que serão selecionadas durante a evolução.
referências e notas
imagem de capa. Conchas Cepaea nemoralis& Cepaea hortensis. André Künzelmann, UFZ.,
Esta fórmula pode ser escrita de uma forma mais convencional como: \pi =\frac{n(n-1))}{2L}\sum _{i=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
Sendo uma amostra de indivíduos em quem o estudou característica foi medido, a variância pode ser definido como a diferença entre a média dos quadrados dos valores e o quadrado da média dos valores. Esta medida, que é sempre positiva, indica a dispersão dos indivíduos.isto é hereditariedade no sentido estrito do termo. A hereditariedade em sentido amplo é a relação entre a soma das variâncias genéticas e a variância total., Esta fórmula dá hereditariedade com ambos os pais (o” pai médio”); a estimativa com apenas um pai daria h2/2.
O Ambiental Enciclopédia do meio Ambiente pela Associação des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contratualmente vinculado à Universidade de Grenoble Alpes e Grenoble INP, e patrocinado pela Academia francesa de Ciências.
to cite this article:VEUILLE Michel (2021), Genetic polymorphism and variation, Encyclopedia of the Environment, url : https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
Os artigos na Enciclopédia do Meio Ambiente são disponibilizados sob os termos da licença Creative Commons BY-NC-SA, que autoriza a reprodução sujeita a: citando a fonte, não fazendo uso comercial deles, compartilhando condições iniciais idênticas, reproduzindo em cada reutilização ou distribuição a menção desta licença Creative Commons BY-NC-SA.
Deixe uma resposta