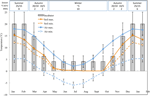
Figura 1. Date lunare maxime și minime de temperatură a aerului și a solului în Alpii Australieni (medie ± STDEV). Temperaturile aerului sunt medii din Baza de date BOM, temperaturile solului au fost măsurate cu iButtons (vezi metode). Barele indică temperaturile experimentale ale incubatorului (media ± STDEV)., Timpul echivalent al anului (sezon), regimul temperaturii incubatorului (°c, 12/12 h, lumină/întuneric) și durata tratamentului (săptămâni), așa cum este indicat în panoul superior.
Cu scopul de a descoperi condițiile de temperatură apropiate de cele care semințele experiența post-dispersie, zilnice maxime și minime de temperatură din sol au fost colectate în cadrul studiului site-ul (de Marinar Colibă; 36.27°S și 148.17°E, 2030 m un.s.l.) folosind ibutton furnizori de bustean de date (n = 10, Încorporate Sisteme de Date, statele UNITE ale americii) plasate 4 cm sub suprafața solului de la baza de vegetație de 17 ianuarie-17 decembrie 2012., Am plasat-furnizori de bustean la 4 cm, pentru a evita perturbare suprafață, dar încă să fie reprezentative pentru condițiile în care semințele sunt expuse în sol; această adâncime este intermediare de la ce a fost folosit în prealabil alpine temperatura solului exerciții de monitorizare (Scherrer și Körner, 2011; Pauli et al., 2015). După cum se bănuia, temperatura medie a solului nu a scăzut sub nivelul de îngheț în timpul iernii (Figura 1). În timpul verii, temperatura solului sub vegetație este cunoscută pentru a urmări temperatura ambiantă, în timp ce temperaturile solului gol vor depăși mediul înconjurător cu până la 15°C (Soil Conservation Service unpublished records, 1960s–1970s).,semințele Mature din 54 de specii din 16 familii și 37 de genuri au fost colectate între ianuarie și aprilie 2009, 2010 și 2011 (a se vedea tabelul 1 pentru numele și autoritățile complete. Voucherele au fost depuse la Herbarul Național Australian, Canberra). În total, specia a reprezentat mai mult de un sfert din flora angiospermă australiană Găsită în regiunile alpine (Costin et al., 2000, vezi Tabelul 1), deși multe dintre aceste specii se extind și sub copac., Viabilitatea tuturor colecțiilor a fost estimată înainte de însămânțare în condiții de germinație experimentală folosind tehnica de colorare a clorurii de tetrazoliu (TZ) (International Seed Testing Association, 2003). Pentru mai multe detalii privind colectarea și prelucrarea, a se vedea anexa A în materialul suplimentar.
Tabelul 1. Studiu de specii, de familie și de autoritate, forma de creștere, și colecții detalii, procentul de semințe de viabilitate (TZ-estimat, ± s. e.), procentul final de germinare realizat (± î,), diferența statistică între viabilitate și germinare (G < V, testată numai în cazurile în care germinarea a fost mai mică decât viabilitatea).fenologia germinării semințelor a fost investigată prin imitarea regimurilor de temperatură pe care semințele alpine le experimentează in situ, post-dispersare, într-o evoluție artificială a anotimpurilor. Semințele au fost îmbibate pe tot parcursul experimentului, astfel rezultatele indică potențialul de germinare atunci când apa nu se limitează., Datele privind temperatura aerului și a solului (Figura 1) au fost utilizate pentru a ghida și valida regimurile de temperatură ale incubatoarelor, deși dezavantajele logistice ale lucrului cu Incubatoarele au împiedicat încorporarea fluctuațiilor inerente condițiilor naturale. Testele de germinare au folosit opt replici de 25 de semințe pe colecție, semănate în vase Petri din plastic cu diametrul de 9 cm care conțin 1% apă simplă-agar. Plăcile Petri au fost sigilate cu Parafilm pentru a evita agar uscare, înainte de a fi introduse în germinare incubatoare (Thermoline Științifice, Melbourne, VICTORIA, Australia)., Jumătate din vase au fost învelite în folie de aluminiu pentru a exclude lumina și pentru a oferi o indicație a cerințelor de lumină pentru germinare. Fiecare dintre cele patru replici per tratament cu lumină per colecție a fost plasat pe un raft diferit în același incubator și locația din acel raft a fost rearanjată săptămânal. Toate repetări din fiecare colecție au fost mutate prin următoarele simulat thermoperiods: 8 săptămâni la 25/15°C (temperatura zi/noapte) → 2 săptămâni de la 20/10°C → 2 săptămâni de la 10/5°C → 10 săptămâni de la constanta 4-5°C → 2 săptămâni de la 10/5°C → 2 săptămâni de la 20/10°C → 6 săptămâni de la 25/15°C., Astfel, un ciclu al întregului experiment a imitat temperaturi care amintesc de vară, începutul toamnei, toamna târzie, iarna, primăvara devreme, primăvara târzie și vara și a durat în total 32 de săptămâni. Anul abreviat a fost considerat rezonabil, deoarece aderările au încetat, în general, germinarea ulterioară într-un anumit tratament înainte de a fi mutate în următorul „sezon”., Un 12/12 h lumină/întuneric iluminare a fost oferit de-a lungul cu tuburi fluorescente (> 90 µmol m−2 s−1 în centrul fiecărui raft), și în primăvara, vara și toamna regimuri de temperatură lumină a coincis cu temperatură caldă perioadă a zilei. Germinarea, definită ca apariția radiculei cu mai mult de 1 mm, a fost marcată la fiecare 7 zile, iar semințele germinate și răsadurile au fost ulterior eliminate din vase. Germinarea plăcilor învelite în folie a fost evaluată ca mai sus, dar sub lumină verde foarte scăzută.,după un ciclu experimental (32 săptămâni), majoritatea colecțiilor au fost terminate (Vezi informații suplimentare pentru excepții), prezentând germinare zero timp de cel puțin 2 săptămâni în acest moment. După terminare, toate semințele intacte rămase au fost disecate cu un bisturiu sub microscop. Semințele cu un endosperm ferm, proaspăt și embrion au fost considerate viabile, iar semințele goale dintr-un embrion au fost deduse din total la calcularea procentului de germinare pentru fiecare colecție., Am calculat un indice al cerințelor de lumină pentru germinare pentru fiecare specie ca fracțiune de germinare în tratamentul întunecat în momentul în care s-au atins percentilele de germinare cumulate de 50 și 95% în tratamentul alternativ lumină/întuneric (vezi mai jos).
corelații ecologice ale Strategiei de germinare
am compilat o bază de date a trăsăturilor funcționale și a măsurilor geografice ale speciilor din datele publicate pentru a evalua corelațiile cu strategia de germinare., Baza de date a inclus următoarele trăsături continue: masa medie a semințelor, cerința ușoară pentru germinare (a se vedea mai sus), suprafața specifică a frunzelor (suprafața proaspătă/masa uscată), suprafața individuală a frunzelor, greutatea uscată a frunzelor individuale și înălțimea la maturitate. Creșterile minime, medii și maxime de colectare pentru specii au fost determinate din toate înregistrările herbarului din Herbarul Virtual Australian, iar elevația la care au fost făcute colecțiile specifice de semințe utilizate în studiu a fost, de asemenea, inclusă ca trăsături continue. Forma de creștere a fost marcată ca una din cele trei categorii: graminoid, plante medicinale sau arbust., Am inclus, de asemenea, o trăsătură binară: dacă semințele erau endospermice sau nu.
trasatura de date au fost colectate de la o varietate de surse, inclusiv Australian Virtual Ierbar (http://avh.chah.org.au/), la Australian National Ierbar specimen informații registru (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Flora Alpină (Costin et al., 2000), Flora din New South Wales (Harden, 1990) și morfologia internă comparativă a semințelor (Martin, 1946). Masa semințelor și suprafața specifică a frunzelor au fost obținute din colecții individuale de teren utilizate în studiile de germinare., Masa semințelor a fost obținută prin cântărirea a patru loturi de 10 semințe. SLA a fost determinată pe baza a cinci frunze complet extinse de la indivizi separați pe specie. Acestea au fost colectate în teren, scanat cu un scaner, uscate la 60°C la greutate constantă și ponderate la cel mai apropiat 0.000 g. În cazul în care insuficientă de semințe a fost disponibil, sau specifice frunze zonă nu a fost disponibil, datele au fost obținute de la alte ANBG colecții din aceeași specie sau din alte surse de date (S. Venn, date nepublicate)., Dacă este sau nu o sămânță a fost endospermic a fost determinată de desene de semințe în ANGB bancă de semințe de înregistrări în consultare cu ANBG semințe de personalul băncii. Baza de date completă și sursele sunt disponibile la cerere de la autori.
analiza datelor
modelele de germinare au fost evaluate vizual și folosind o analiză cluster în PC-ORD Pentru Windows, versiunea 6 (MJM Software Design, Gleneden Beach, sau, SUA). Analiza a folosit o matrice de distanță euclidiană și un algoritm de îmbinare cel mai apropiat vecin., Punctele de date pentru analiza cluster au fost timpul (săptămâni) până la percentilele de germinare cumulative specifice (t20, t40, t50, t60, t80 și t95%) pentru fiecare colecție (medie pe cele 4 feluri de mâncare). Astfel, o colecție care a germinat la începutul experimentului ar avea perioade scurte până la percentile până la t80%, de exemplu, în timp ce o colecție care a amânat germinarea până când temperaturile au crescut în urma iernii simulate ar avea perioade lungi până la aceste percentile., Mai complexe de germinare modele, cum ar fi lent, constant germinare sau exploziile de germinare, de asemenea, ar putea fi detectat, și ar fi indicat de uniform distanțate ori sau ori grupate la începutul și la sfârșitul experimentului, respectiv. Această metodă a fost utilizată ca o modalitate de a impune o evaluare obiectivă a modelului de germinare, independent de temperatura în sine.,o analiză unidirecțională a varianței (ANOVA) a fost efectuată pentru plăcile reproduse ale fiecărei colecții pentru a evalua dacă a existat o diferență semnificativă între viabilitatea procentuală (determinată prin testul TZ) și Germinarea procentuală finală. Alte modele liniare generale au fost utilizate pentru a evalua dacă clusterele rezultate din analiza clusterului diferă în ceea ce privește viabilitatea sau procentul final de germinare.asociațiile dintre trăsăturile ecologice și grupurile de strategie de germinare au fost testate în două moduri., În primul rând, am testat diferențele medii între clustere folosind ANOVA cu cluster ca efect fix și corelate ecologice ca variabile de răspuns sau folosind teste g replicate pentru endospermie și formă de creștere. În al doilea rând, deoarece germinația și alte trăsături funcționale sunt foarte susceptibile de a fi influențate de istoria evolutivă, am evaluat aceste corelații folosind regresii filogenetice. A fost construită o filogenie pentru speciile de studiu folosind Filocom Filomatic și ambalajul bladj în R (Webb et al., 2008; Echipa De Bază R, 2013)., Am evaluat prezența structurii filogenetice în variabile folosind filosig sau filo.d (variabile discrete sau binare) funcția pachetului Fitools în R (Revell, 2012). Regresiile filogenetice au fost efectuate utilizând funcția pgls în Caper (Orme et al., 2013). Analizele au fost efectuate într-un mod structurat, reflectând gruparea evidențiată în PC-Ord (vezi mai jos)., Inițial am inclus patru strategii de germinare (amânate, imediate și eșalonate în fiecare dintre cele precedente), apoi trei (lumping toate speciile eșalonate și comparând cu amânate și intermediare) și în cele din urmă am evaluat diferențele dintre cele două strategii mai largi, amânate și intermediare.
rezultate
viabilitatea semințelor comparativ cu germinarea finală
În general, viabilitatea semințelor și procentul de germinare finală au fost ridicate. Viabilitatea medie estimată de TZ pentru toate colecțiile a fost de 80 ± 1.,7%, cu mai mult de două treimi din colecții care prezintă viabilitate mai mare de 75% (Tabelul 1). Nu a fost nici o diferență în calitatea de colecții de peste 3 ani (adică viabilitatea în 2009, 2010 și 2011 a fost de 79 ± 2.8, 80 ± 2.7, și 81 ± 3,7%, respectiv), indicând faptul că sistemul bancar de semințe colectate în anul 2009 nu a reduce viabilitatea acestora.
Germinare care a depășit sau nu a fost semnificativ mai mic decât TZ-estimat viabilitatea a fost realizat pentru 45 din 54 de specii (One-Way ANOVA: P>0.05, Tabelul 1)., Dintre cele nouă specii rămase, germinarea finală a patru specii a fost foarte scăzută (< 5%), în ciuda viabilității bune (ca. 55-94%) și astfel nu am putut trage concluzii cu privire la strategiile de germinare ale acestor specii. Eșecul de a germina ar putea reflecta fie că tratamentele noastre nu au atenuat starea de dormit, fie că semințele nu au fost pe deplin mature la colectare. Testele tăiate la sfârșitul experimentului au indicat că, în toate cazurile, majoritatea semințelor ne-germinate conțineau un embrion sănătos, îmbibat., Interesant este că 34 de specii au obținut o germinare mai mare decât viabilitatea estimată TZ (Tabelul 1), sugerând că testul TZ a subestimat în mod obișnuit viabilitatea semințelor.
clustere de germinare
analiza clusterelor a relevat două clustere majore (A și B), cu sub-structură în fiecare (figura 2). Pentru majoritatea speciilor (n = 21 specii din 10 familii), germinarea a fost amânată până într-o perioadă de temperaturi de iarnă (5°C zi/noapte) sau până când temperaturile au fost ridicate la 10/5°c după iarnă (A1 și A1:iarnă, figurile 2, 3a)., Alternativ, germinarea a început imediat la 25/15°c (n = 17 specii, șase familii, observate în clusterul B1, figurile 2, 3b). Pentru majoritatea acestor specii, rata de germinare a fost rapidă; 50% din germinația totală (t50) a avut loc în mai puțin de 3 săptămâni (figura suplimentară 1).
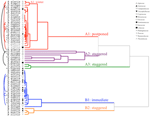
Figura 2. Analiza Cluster dendrogram bazat pe timp la percentilele germinare cumulative(20, 40, 50, 60, 80, și 95%). Specii indicate prin genul inițial și prima parte a numelui speciei. Asteriscurile marchează colecțiile care au fost bancate înainte de testare., Arcele Fine conectează colecțiile duplicate pentru a compara efectul băncilor asupra germinării. Simbolurile denotă diferite familii (vezi legenda). Colecțiile prezentate în text gri sunt cele care au fost expuse la două cicluri ale experimentului.
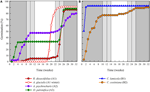
Figura 3. Germinarea procentuală cumulativă a unei specii reprezentative din fiecare grup de germinare. (A) grupa A, specii care au amânat germinarea, (B) Grupa B, specii care au germinat imediat., Rețineți că modelele de germinare eșalonate sunt imbricate în grupurile de mai sus. Regimurile de temperatură ale incubatorului sunt reprezentate prin umbrire (a se vedea Figura 1).în cadrul fiecăreia dintre aceste clase largi, cu toate acestea, au existat sub-clustere clare care reflectă germinarea care a fost eșalonată în timp, adică o proporție din lotul de semințe a germinat înainte de expunerea la temperaturile de iarnă, în timp ce restul a germinat după iarnă. Germinarea eșalonată a avut loc la 13 specii din șase familii și a avut loc în principal atunci când temperaturile din timpul zilei au depășit 10°C (figurile 3a,B)., Acești germinatori eșalonați au fost împărțiți în trei grupuri în analiza clusterului. Clusterul A2 cu șapte colecții a arătat o germinare substanțială atât înainte, cât și după perioada de temperatură rece. În cazul în care germinarea a încetat înainte de schimbarea temperaturilor, am concluzionat că lipsa germinării ulterioare înainte de iarnă nu a fost determinată de temperatură (A3). În schimb, germinarea unei mari părți a grupului A2 părea să fie oprită numai atunci când temperaturile au fost reduse la 10/5°C., Clusterul B2 conținea încă patru specii care au prezentat o germinație substanțială la începutul experimentului și au obținut o proporție relativ mică din germinația lor totală după ce temperaturile au revenit la peste 5°C (figura 3b).comparativ cu variația relativ minoră dintre speciile din grupurile A1 și B1, grupurile eșalonate prezintă, de asemenea, o diferențiere mult mai mare între specii. Curbele individuale pentru toate speciile sunt prezentate în figurile suplimentare 1i-vii. pentru detalii mai specifice privind strategia de germinare, a se vedea apendicele B din materialul suplimentar.,dintre toate corelațiile ecologice pe care le-am considerat, doar cele asociate direct cu trăsăturile semințelor au fost corelate semnificativ cu strategia de germinare indiferent dacă am evaluat corelația la nivelul a două clustere majore (amânate vs.germinarea imediată), sau la nivelul a trei sau patru clustere, sau dacă am contabilizat structura filogenetică sau nu (Tabelul 2)., Speciile cu germinare amânată au avut semințe mai grele și au fost mai susceptibile de a fi endospermice în comparație cu cele ale speciilor care au germinat imediat (Figura 4). Speciile cu germinare eșalonată au fost intermediare în masa semințelor și endospermie. Cu toate acestea, nu toate caracteristicile semințelor au fost corelate cu strategia de germinare: nu a existat o corelație între cerințele ușoare pentru germinarea semințelor și strategia. Contrar așteptărilor noastre, speciile cu o suprafață specifică mai mare a frunzelor (potențial indicatoare ale unor rate de creștere mai mari) nu aveau mai multe șanse să prezinte germinare imediată., De asemenea, speciile cu altitudine medie mai mare sau intervale de altitudine mai mici nu au fost mai susceptibile să prezinte germinare amânată.

Tabelul 2. Incidența filogenetic structură, și se corelează de germinare strategie.
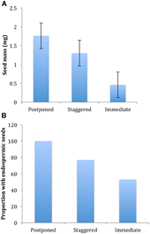
Figura 4., Diferențele dintre (a) masa medie a semințelor și (B) proporția speciilor cu semințe endospermice între speciile cu strategii amânate (clusterul A1), eșalonate (combinația dintre clusterul A2, A3 și B2) și Germinarea imediată (clusterul B1). Diferențele dintre strategii sunt semnificative la p ≤ 0,05, a se vedea tabelul 2.
Discuții
Germinare în alpine habitatelor din punct de vedere istoric a fost considerat a fi mediu controlat, cu zăpadă iarna izolator semințe împotriva potențialelor germinare indicii și de redare hibernare inutile (Billings și Mooney, 1968)., Cu toate acestea, rezultatele noastre susțin dovezi mai recente ale faptului că mecanismele de dormit sunt comune în rândul speciilor alpine atât în Australia (Hoyle et al., 2013; Sommerville și colab., 2013) și în altă parte în jurul lumii (Cavieres și Arroyo, 2000; Schutz, 2002; Shimono și Kudo, 2005; Mondoni et al., 2009; Schwienbacher și colab., 2011)., Hibernare pare să joace un rol semnificativ în controlul in situ calendarul de germinare de multe Australian specii alpine, hotărând să amâne, să amâne, sau încetini rata de germinare și, astfel, potențial care conferă aversiune la risc de regenerare strategii într-un mod dur și variabile de mediu. Poate cel mai frapant al nostru germinare rezultate fost proporția de specii pentru care germinare strategii variate în cadrul unei colectare a semințelor pentru o anumită specie sugerând că ambele latente și non-latente caracteristici au fost expuse în cadrul aceleași semințe de colectare a acestor specii., În special, strategiile de germinare au fost în general foarte conservate în cadrul familiilor și, de asemenea, corelate cu elemente ale anatomiei semințelor: masa și endospermul. Strategiile de germinare au fost, totuși, independente de alte trăsături funcționale semnificative din punct de vedere ecologic ale plantei mature.
variația Strategiei de germinare
semințele a aproape jumătate din speciile studiate au părut incapabile să germineze până când nu au fost expuse la o perioadă rece și umedă (constantă 5°C), sugerând că stratificarea la rece a atenuat un mecanism fiziologic de dormit., Amânarea germinării până în primăvara următoare poate permite răsadurilor să evite instalarea peste sau înainte de iarna aspră, optimizând în același timp sezonul scurt de creștere care urmează. În schimb, germinarea oportunistă expusă de speciile care au germinat imediat poate oferi un avantaj selectiv atunci când riscul mortalității răsadurilor de iarnă este scăzut, permițând plantelor să înflorească mai devreme în primăvara următoare sau la o dimensiune mai mare (Donohue, 2002).,până în prezent, au existat puține dovezi publicate despre variabilitatea intra-specifică a mecanismelor de dormit în cadrul colecțiilor de semințe alpine, cum ar explica strategiile de germinare eșalonate observate aici. Germinarea eșalonată în timp poate fi, de asemenea, explicată prin niveluri diferite de maturitate a semințelor în rândul indivizilor din populație la momentul colectării și/sau ar putea reflecta o strategie dimorfică într-o singură plantă asociată cu poziția pe plantă sau cu calendarul de dezvoltare.,
evolutive bet-hedging este adesea evocat pentru a explica strategia diversificată în caracteristicile de germinare a semințelor (Simons, 2011), cum ar fi cea demonstrată de colecțiile cu germinare eșalonată. Dacă, totuși, o gama diversificata de strategie rezultate în ambele crescut medie de fitness și o reducere a varianței (de exemplu, prin reducerea concurenței între frați), atunci aceasta nu constituie bet-de acoperire a riscului. De asemenea, dacă aparent diversificarea reflectă diferențele dintre, nu se încadrează, persoane fizice, atunci aceasta nu este bet-acoperire (Starrfelt și Zamor, 2012)., Investigațiile alpine răsad de dispersie modele și toleranță de îngheț și zăpadă se topesc, asociat cu studii de factori determinanți de germinare strategia de variație în cadrul speciilor, ar putea explica de germinare fenologia speciilor care prezintă un eșalonate strategie și poate indica dacă acestea sunt susceptibile de a reprezenta bet-strategii de acoperire.,
corelații ecologice ale Strategiei de germinare
examinarea relației dintre strategia de germinare și alte trăsături importante din punct de vedere ecologic a demonstrat că trăsăturile de germinare au fost corelate cu alte trăsături ale semințelor, dar nu și cu cele vegetative. Sommerville și colab. (2013) a constatat că ierburile Alpine australiene cu semințe non-endospermice au fost mai susceptibile de a nu fi latente la dispersare. Munca noastră susține această constatare și indică în continuare că semințele mai mici sunt mai susceptibile de a fi printre germinatorii imediați. Deși Sommerville și colab., (2013) nu a posit de ce endospermia poate fi asociată cu latența, sugerăm că plasarea rezervelor în cotiledoane poate îmbunătăți rata de creștere timpurie și stabilirea în speciile germinative imediate. Rezultatele noastre nu au indicat nicio corelație între cerința de lumină și starea de repaus, deși cerințele de lumină variază între specii. Există o varietate de alte trăsături de semințe care tamponează riscul de dispariție în medii variabile, de exemplu mecanismele de dispersie a semințelor și longevitatea semințelor în banca de semințe de sol (vezi tielborger et al., 2012 pentru o revizuire)., În acest stadiu nu putem spune dacă aceste trăsături ar arăta corelații cu strategia de germinare sau nu.
am întrebat în continuare dacă strategia de germinare ar fi corelată cu trăsături non-reproductive importante din punct de vedere ecologic, cum ar fi zona specifică a frunzelor sau înălțimea plantelor (Westoby, 1998; Wright et al., 2004). În special, am afirmat că speciile germinative imediate ar putea prezenta valori ale trăsăturilor care indică rate de creștere mai rapide (folosind SLA ca proxy), așa cum ar putea fi necesare pentru stabilirea în perioada scurtă dintre dispersare și debutul iernii., Cu toate acestea, nu am găsit astfel de corelații. Dacă trăsăturile fiziologice și de creștere ale răsadurilor în stadiile timpurii de înființare ar putea prezenta modele diferite decât trăsăturile vegetative adulte, este o întrebare demnă de luat în considerare, dar dincolo de domeniul de aplicare al prezentei lucrări.
în cele din urmă, am examinat dacă speciile de la altitudini mai mari, unde anotimpurile de creștere sunt în general mai scurte, ar putea fi mai susceptibile de a prezenta latență, dar nu a existat nicio asociere între intervalul de altitudine, creșterile de colectare sau creșterea medie a colectării pentru specia în ansamblu și strategia de germinare., Împreună, aceste rezultate indică faptul că, deși Evoluția strategiei de germinare este strâns legată de trăsăturile importante din punct de vedere ecologic ale semințelor, ea cuprinde o axă în mare măsură independentă față de caracteristicile vegetative sau distributive.implicațiile pentru comunitățile de plante Alpine într-un climat în schimbare ecosistemele Alpine australiene, cum ar fi zonele alpine din întreaga lume, sunt amenințate de schimbările climatice combinate cu schimbări în frecvența incendiilor, modelele de utilizare a terenurilor, afluxul de specii invazive și impactul vizitării umane crescute., Încălzirea asociată cu schimbările climatice se produce mai rapid deasupra liniei copacilor decât la altitudini mai mici, iar zonele alpine vor continua să experimenteze încălzirea peste medie în viitor (Kullman, 2004). Capacitatea de regenerare continuă prin semințe în condiții noi este probabil să joace un rol semnificativ în răspunsul plantelor alpine la schimbările climatice și la schimbările asociate. În plus, este posibil ca regimurile de perturbare modificate să necesite un rol sporit de restaurare și reabilitare în regiune., Informațiile furnizate aici sunt, prin urmare, crucială pentru conservarea și gestionarea alpine sistemelor în curs de schimbare; ele pot ajuta la informarea previziunile noastre de cât de studiu specia poate fi afectată, formează baza de semințe de propagare planuri pentru aceste specii, și să fie utilizate pentru a ghida viitorul ancheta alpine germinarea semințelor strategii cu identificabile de conservare și de gestionare a rezultatelor.,schimbările de temperatură pot afecta starea de latență a semințelor atât pre-cât și post-dispersare, modificând și/sau perturbând strategiile de germinare și ducând la schimbări în fenologia germinației curente (Hoyle et al., 2008b). Pentru plantele alpine, încălzirea climatică poate duce la o schimbare de la apariția primăverii la apariția toamnei, determinată în primul rând de modificările stării de inactivitate a semințelor și având ca rezultat implicații majore pentru speciile adaptate în prezent la apariția primăverii (Mondoni et al., 2012)., În continuare cercetări în modul în care schimbările la temperaturi că semințele experiența post-dispersie, a redus durata stratului de zăpadă, și a crescut frecvența de temperaturile extreme vor afecta germinare sunt necesare în scopul de a prezice cât de specii cu latență va răspunde la schimbările climatice. Pentru acele specii alpine care germinează în toamnă, înainte de îngheț și zăpadă, rezultatele noastre ridică problema modului în care răsadurile acestor specii vor face față reducerii preconizate a precipitațiilor și frecvenței crescute a evenimentelor de îngheț sezonier.,în cele din urmă, rezultatele noastre ridică întrebări importante cu privire la variația în cadrul speciilor, în special în asociere cu prezicerea impactului schimbărilor climatice sau selectarea semințelor pe care să le utilizeze pentru restaurare. Strategiile de germinare pot varia în funcție de specie, în funcție de populație, timpul de înflorire, masa semințelor, investiția parentală, clima, elevația și/sau mediul matern, iar această variație poate reflecta factorii genetici, epigenetici și/sau de mediu (Cochrane et al., 2015)., Creșterea altitudine conturile de variație în germinability și/sau starea de hibernare de mai multe specii alpine (Billings și Mooney, 1968; Dorne, 1981; Cavieres și Arroyo, 2000), dar vezi (Hoyle et al., 2014). Starea redusă de inactivitate poate rezulta din încetarea anticipată a dezvoltării semințelor care rezultă din stresul de mediu (Steadman et al., 2006; Hoyle și colab., 2008b)., Examinarea variației strategiilor de germinare în cadrul speciilor poate dezvălui potențialul speciilor de a face față și de a se adapta la schimbările climatice, jucând astfel un rol important în determinarea supraviețuirii viitoare și a distribuțiilor speciilor și oferind o perspectivă importantă pentru gestionare și conservare.abordarea noastră a relevat apariția pe scară largă a latenței în flora alpină australiană și demonstrează că strategiile de germinare variază atât în cadrul unei colecții de semințe, cât și între colecții., Mai mult, am arătat că, în timp ce strategia de latență este conservată filogenetic și corelată cu masa semințelor și endospermia, este în mare măsură independentă de trăsăturile vegetative și caracteristicile gamei. Deși nu se poate extrapola direct din rezultatele noastre modul în care schimbările climatice și asociate vor afecta flora alpină, aceste rezultate pot constitui baza pentru proiectarea planurilor de propagare și experimentarea ulterioară a impactului regimurilor climatice în schimbare., În special, susținem cercetări suplimentare în înțelegerea cauzelor variației în cadrul speciilor în strategia de germinare și a rolului pe care acesta îl poate juca în capacitatea speciilor de a rezista schimbărilor climatice preconizate.
accesibilitatea datelor
datele complete de germinare și baza de date a trăsăturilor vor fi depuse pe DRYAD și cu Atlasul Australiei vii.
Declarație privind conflictul de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.,
mulțumiri
recunoaștem finanțarea de la Consiliul Australian de cercetare (Lp0991593) și Grădina Botanică Națională Australiană și prietenii acestora. Multumim lui J. McAuliffe, S. Fethers si S. Lawatsch de la ANBG. De asemenea, mulțumim M. Holloway-Phillips, M. Cardillo, și S. Cunningham pentru intrare pe versiunile anterioare ale manuscrisului și pentru sfaturi analitice și S. Venn pentru accesul la date trăsătură.
material suplimentar
Foley, me (1992). Efectul zaharurilor solubile și al acidului giberelic în ruperea latenței embrionilor excizați de ovaz sălbatic (Avena fatua)., Weed Sci. 40, 208–214.Google Scholar
Harden, GJ (1990). Flora din New South Wales. Sydney, NSW: Royal Botanic Gardens.Google Scholar
Asociația Internațională de testare a semințelor. (2003). Fișe de lucru pentru testarea Tetrazoliului. Bassersdorf: ISTA.Kullman, L. (2004). Fața schimbătoare a lumii alpine. Glob. Schimba Newsl. 57, 12–14.
Google Scholar
Lasă un răspuns