
polimorfismul Genetic este existența unor alternative membre ale ADN-ului, determinarea variației organismului grad mai mare de integrare. Există diferite tipuri de modificări ale genomului (mutații). Cele mai studiate sunt substituțiile nucleotidice în regiunile de codificare și reglementare.
Definitie
polimorfismul Genetic este existența, într-o populație, din mai multe alternative membre ale ADN-ului, sau alele, la o anumită poziție în genom, sau locusPosition a genei pe cromozom., În genetica populației, un set de gene omoloage (clasa omologie). Două gene sunt omoloage dacă se potrivesc cu meioza. Plural: loci.. Această definiție are mai multe aspecte:
(1) în primul rând, trăsătura trebuie să fie purtată de cromozomi și să fie transmisibilă.
(2) atunci alelele trebuie să fie omologedouă cromozomi sau două gene se spune că sunt omoloage dacă se potrivesc și se exclud reciproc de meioză. pentru poziția lor în genom, care exclude anumite tipuri de variații care vor fi discutate mai târziu.,
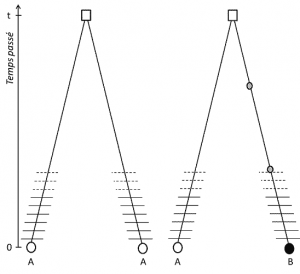
(3) dar, deoarece trăsătura este transmisibilă, omologia poziției implică, de asemenea, că alelele sunt omoloage prin descendență; dacă sunt diferite, prin urmare, a apărut o mutație (cel puțin) într-una din liniile care le leagă de ultimul lor strămoș comun (Figura 1).
(4) în cele din urmă, polimorfismul genetic poate fi definit la scara celei mai mici unități care compune ADN: situl nucleotidic., Ca rezultat, fiecare variantă nucleotidică poate determina polimorfismul nivelurilor superioare ale structurii biologice – gena, proteina și fenotipul individului-care devin Scale la care poate fi descris polimorfismul genetic (Figura 2).luați exemplul sistemului sanguin ABO, care corespunde motivelor antigeniceo parte a antigenului recunoscut de un anticorp sau de un receptor de limfocite. De asemenea, cunoscut ca un epitop sau determinant antigenic. Același antigen poate avea mai mulți epitopi (identici sau diferiți) și astfel induce un răspuns imun variat., implicat în Compatibilitatea în timpul transfuziilor de sânge. Acesta este un caz de polimorfism genetic prezent în populațiile umane, implicând proteina ABO glicozil transferazeenzima care permite transferul reziduurilor care conțin zaharuri în proteine. În cazul sistemului ABO, glicozil transferaza A și, respectiv, B induc unui individ să aparțină grupei A sau B. Când sunt prezente ambele glicoziltransferaze, individul este din grupa AB. care are trei alele, A, B și O. un individ poate avea genotipuri (AA), (AO); (BB), (BO); (oo) sau (AB)., Fenotipul său va fi atunci,,;,; sau, unde vedem că A și B „domină” O (adică expresia lor o maschează pe cea a lui O) și că sunt co-dominante între ele (heterozigotul dintre A și B are o expresie recognoscibilă ).un polimorfism poate fi descris mai întâi la scara secvenței ADN a locusului care codifică o proteină. Unele polimorfisme sunt „sinonime”, adică ele nu schimbă secvența de aminoacizi a proteinei; ele sunt adesea cele mai numeroase polimorfisme. Ceilalți modifică aminoacizii și se numesc polimorfisme „de înlocuire”., Polimorfismele de înlocuire ale glicozil transferazei ABO sunt de două tipuri: polimorfisme care schimbă aminoacidul, dar nu unitatea antigenică (nu interferează cu fenotipul ABO al indivizilor); și alte polimorfisme de înlocuire, care determină fenotipul ABO al caracteristicilor observabile ale unui individ..
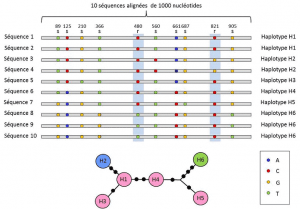
Între nucleotide și alela niveluri, analize genetice ia în considerare un nivel intermediar descriere: aranjamentul liniar variabilă site-uri de pe locus. Aceste aranjamente, numite haplotipegrup de alele de diferite loci situate pe același cromozom și, de obicei, transmise împreună. Haplotipul este format prin contracția genotipului haploid al expresiei engleze sau a genotipului haploid., Toate genele situate pe același cromozom și ale căror alele se separă împreună în timpul meiozei constituie un haplotip. Aceste gene sunt numite „legate genetic”, sunt utile în genetica evolutivă, deoarece permit găsirea legăturilor genealogice dintre alele (Figura 2).polimorfismele nucleotidice există, de asemenea, în regiunile intergenice (în afara regiunilor de codificare). Unele afectează zonele în care expresia genelor este reglementată și, prin urmare, au Expresie fenotipică. Altele nu au efect cunoscut și sunt denumite „silențioase”.,
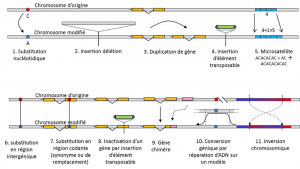
În plus față de substituții nucleotidice, polimorfism pentru un anumit tip de mutație, microsateliți, se referă la variații în lungimea de multe ori non-codare fragment de ADN (Figura 3) din cauza la o schimbare în numărul de repetiții într-un scurt repetă secvența de nucleotide, de exemplu, CACACACA, sau TGTGTG.nu toate variațiile genetice se încadrează în definiția polimorfismului genetic, deoarece necesită omologia poziției alternativelor (Figura 3)., Astfel, există secvențe repetate obținute prin duplicarea unei gene, pentru care două repetări nu sunt omoloage pe poziție. Adesea, nu putem vorbi nici despre omologie pentru elementele transpozabile, deoarece în general își schimbă poziția în timpul generațiilor celulare, se pot multiplica și invada genomul. Omologia a două elemente transpozabile nu poate fi apoi definită. Pe de altă parte, două secvențe de codificare ale aceluiași locus, dintre care unul este intact, iar celălalt inactivat prin inserarea unui element transpozabil, corespund bine unei situații de omologie., Într-o zi, vom lua în considerare probabil să vorbim despre polimorfism pentru modificările așa-numitelor cromozomi „epigenetici”, care sunt uneori transmise pe mai multe generații celulare, somaticcalifică celulele non-reproductive sau soma. O mutație somatică, care afectează o genă a unei celule somatice, dispare cu individul purtător. sau germinalQualifies gameți. O mutație a liniei germinale poate fi transmisă descendenților săi., cum ar fi metilarea. Importanța lor în evoluția populațiilor rămâne de evaluat., Limitarea noțiunii de polimorfism genetic la anumite categorii de variație – în esență substituții de nucleotide – provine din Marea utilitate a acesteia din urmă în explorarea istoriei populațiilor naturale. Ele se pretează la modelarea matematică a evoluției.cuvântul polimorfism înseamnă „mai multe forme”. Se opune monomorfismului, care indică absența variației. În vocabularul biologiei, această opoziție a fost folosită de mult timp, înainte de genetică, pentru a semnala coexistența mai multor tipuri de indivizi din aceeași specie, de exemplu., polimorfismul castei (Regina și lucrătorii) la insectele sociale; polimorfismul sezonier (modificări ale blănii) la unele mamifere arctice. Aceste cazuri nu se încadrează în polimorfismul genetic, un concept mai recent, cu un sens mai restrâns și mai precis. Variațiile continue (de exemplu, variația dimensiunii) nu intră, de asemenea, în categoria polimorfismului, deoarece nu prezintă alternative distincte. Cu toate acestea, locus care afectează dimensiunea se încadrează în această definiție., Vom discuta mai jos cele mai studiate variații genetice deoarece acestea sunt cele mai importante pentru evoluție, substituțiile de nucleotide, iar apoi vom discuta variația fenotipurilor.nu este suficient să spunem că un locus este mai mult sau mai puțin variabil fără a da o estimare cantitativă acestei judecăți. Un cercetător poate studia variația la diferite scări. Dacă este interesat doar de alelele unei proteine, el va măsura diversitatea alelică, simbolizată prin „H”. Dacă este interesat de diversitatea ADN, el va măsura diversitatea nucleotidelor, simbolizată prin „π”.,
alele diversitatea H este definit ca probabilitatea de desen două alele diferite pe parcursul a două eșantionare cu replacementPerforming o succesive trage cu livrare din p token-uri într-o urnă care conține n jetoane, înseamnă a lua un prim semn, citind valoarea acesteia, punând-o înapoi în urnă, având un al doilea semn, citind valoarea acesteia, punând-o înapoi în urnă, etc., până la PTH token. Aceasta înseamnă alegerea obiectelor p între n cu repetiție (puteți alege același obiect de mai multe ori) și în ordine (ordinea în care alegeți obiectele este importantă)., Numărul de remize succesive cu jetoane între n este: n × n × n × … × n = NP.. Dacă numim pi frecvența alelei de rangul i, arătăm că probabilitatea de a alege de două ori aceeași alelă este F = Σ pi2. Diversitatea alelică a probei este apoi complementul său la 1, adică:
H = 1 – Σ pi2 (1)
aceasta se aplică atât alelelor proteice, cât și haplotipurilor. Aceasta se numește diversitate haplotipică. Se poate numi și heterozigozitate, deoarece în cazul unui loc diploid, dă frecvența așteptată a heterozigoților.,
diversitatea nucleotidelor π este echivalentul lui H la scara fiecărei nucleotide. Se calculează ca medie a numărului de diferențe de nucleotide între secvențele dintr-o probă prelevată două câte două (δij), împărțită la lungimea fragmentului ADN în numărul de nucleotide (l).
π = medie (δij) /l (2) (vezi referința)
această valoare diferă între specii. În părțile de codificare ale genomului uman, doi cromozomi selectați aleatoriu din populație diferă în medie cu o nucleotidă la mie., În zbura de fructe (Drosophila melanogaster) această diferență este de aproximativ una din o sută. Zbura este astfel de zece ori mai variabilă decât omul. Atunci când aceste valori sunt legate de dimensiunea regiunile de codificare (aproximativ 15.500 de gene la Drosophila, cel puțin 22.000 de la om), sau chiar de genomuri (140 de milioane de perechi de baze per genom haploid în zboara, aproximativ 3.200 de milioane de euro sau de douăzeci de ori mai mult la om), numărul de site-uri polimorfice este astronomic și rezultatele în nici una fiind născut din sexualizat generație în orice specie este, a fost și va fi genetic identic cu un altul din trecut, prezent sau viitor., Puterea informațională a polimorfismului ADN este imensă. În criminalistică, detectivii pot identifica orice suspect din șaisprezece locusuri microsatelite.
Istorie
termenul variație este în titlul primelor două capitole ale cărții lui Darwin Originea speciilor (1859). Darwin a introdus această noțiune în științele naturii într-un moment în care legile eredității biologice au rămas un mister și i-au dedicat o altă carte importantă, variația animalelor și plantelor (1868)., Convins că evoluția a fost de aproximativ variații mici care a făcut o diferență mică de adaptare a organismelor la condițiile lor de viață, el a atașat o importanță crucială pentru mici variații cantitative, ceea ce a determinat succesorii săi (în special Karl Pearson) a găsit biometryScience de viață de măsurare. Se referă într-un sens foarte larg la studiul cantitativ al ființelor vii.. Dar redescoperirea legilor lui Mendel în 1900 a schimbat interesul pentru variații discontinue.
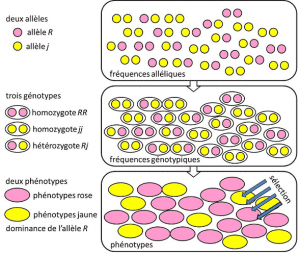
Din 1908 până în 1930, genetica populației a fost în esență o disciplină teoretică care caută să se împace cu Darwinismul Mendeliană heredityHeredity bazate pe transmiterea de o singură genă sub o dominantă, recesivă sau cromozomilor de sex-legate de modul de X (sau Y). Se referă la o trăsătură genetică cu determinism simplu, de un cuplu sau un număr mic de cupluri de gene., și unde probabilitățile au jucat un rol major. Genetica este contra-intuitivă., Ea prezice că nu există o reproducere a părintelui în descendentul său, deoarece genotipul acestuia din urmă rezultă din loteria separării alelelor înainte de fuziunea celor două jumătăți de genom parental. Sa realizat că acestea sunt frecvențele alelicefrecvența la care se găsește alela unei variante într-o populație. Exprimat ca proporție sau procent. Suma frecvențelor alelelor tuturor alelelor unei gene dintr-o populație este, prin urmare, prin definiție egală cu 1. În genetica populației, frecvențele alelelor reprezintă diversitatea genetică la nivel de populație sau specie., care sunt transmise între generații, nu genotipuri sau fenotipuri. Aceste frecvențe sunt mai mult sau mai puțin stabilă de la o generație la alta și de a produce, dincolo de mai multe incurcatura de filiations între indivizi, la fel genotipice frequenciesGenetic structura populației. Determinată din frecvențele alelice (Figura 4). Distribuția genotipurilor populației, asociată cu varianța, este, prin urmare, singurul factor previzibil., În jurul anului 1930, cei trei teoreticieni Ronald Fisher, JBS Haldane și Sewall Wright au ajutat la crearea noțiunii de populație Mendelianăpopulație a cărei ereditate a respectat legile lui Mendel. . În acest context, evoluția asociază în mod necesar trei niveluri structurale: gena, individul și populația. Această asociere cu trei căi poate fi rezumată prin următoarea formulă: evoluția populației este o schimbare a frecvenței alelelor (scala genelor, dar și scara grupului), o schimbare care depinde de sortarea fenotipurilor prin selecție (scala individuală)., Empirice genetica populației de cercetare dezvoltat apoi, dar a fost limitat pentru o lungă perioadă de timp, prin ignoranța noastră de cât de cromozomi de lucru (structura ADN-ului a fost înțeles în 1953 și secvențiere a început în 1977), și a trebuit să cadă înapoi pe studiul de câteva vizibile polimorfisme, cum ar fi colorația gărgăriță elitrele sau coajă de melci (a se vedea Focus marele melc dezbatere). Au existat multe dezbateri între geneticieni cu privire la faptul dacă populațiile naturale erau în general polimorfe sau monomorfe și dacă polimorfismul era în sine benefic sau nu., Studiul variației scării macromoleculare a trebuit să aștepte până în 1966 (pentru proteine) și 1983 (pentru ADN) înainte de a răspunde la aceste întrebări.când ne uităm în jurul nostru, vedem diferențe fizice între ființe, unele complexe precum trăsăturile faciale, altele ușor de măsurat precum greutatea sau înălțimea. Bunul simț sugerează că acestea sunt parțial transmisibile, deși într-un mod dificil de specificat. Aceste trăsături sunt adesea poligenicesub dependența multor gene. Vorbim despre ereditate poligenică. Diabetul este o boală poligenă.,, ceea ce înseamnă că acestea sunt influențate de multe locusuri. Genetica dezvoltării și aplicarea acesteia la compararea speciilor (evo-devo, pentru „evoluția dezvoltării”) dezvăluie rețeaua interacțiunilor complexe care fac ca în timpul dezvoltării celulele corpului, toate cu bagaje genetice identice, să se diferențieze prin epigeneză pentru a da diferite țesuturi. Încă nu este clar cât de complexe sunt construite caracterele. Fenotipurile măsurabile pot fi studiate folosind biometrie, pe baza proprietăților distribuțiilor statistice., O valoare cu proprietăți utile este variantaîntr-un eșantion de indivizi în care a fost măsurată o anumită trăsătură, varianța este diferența dintre media pătratului valorilor și pătratul mediei valorilor. Această măsură, care este întotdeauna pozitivă, indică dispersia indivizilor. . Atunci când mai multe cauze independente determină variația unei linii, variațiile lor sunt aditive și suma lor dă variația liniei. Dacă nu sunt independente, suma covarianțelor se adaugă la suma varianțelor., Varianța totală Vt de o trăsătură fenotipică este astfel încât :
Vt = Vga+Vgd+Vgd+Vgi+Ve
Vga, varianța genetică aditivă, este suma varianțelor datorate pentru fiecare locus în mod independent de ceilalți; Vgd, varianța genetică dominantă, rezultate din interacțiunea dintre cele două alele ale aceluiași locus; Vgi, varianța de interacțiune între locuses, rezultă din faptul că locuses de același individ interacționează; Ve, varianța de mediu, este de presupus că aici independente de variatie genetica.,varianța genetică a dominanței este comună pentru doi copii ai acelorași părinți, deoarece împărtășesc efecte de dominare atunci când la un anumit loc au primit aceleași alele de la ambii părinți, ceea ce li se întâmplă la unul din patru locus. Ei nu împărtășesc aceste efecte cu părinții lor, deci sunt mai „asemănători” între ei decât cu părinții lor, în ciuda faptului că împărtășesc atât de multe gene între ele. Cu siguranță, un părinte poate, din întâmplare, să fi primit aceleași două alele ca unul dintre copiii săi la un anumit locus., Această șansă depinde de frecvența alelelor din populație. Vedem că gradul de similitudine între un copil și părintele său pentru o anumită trăsătură este o proprietate publică.în general, toate componentele varianței genetice (Vga+Vgd + Vgi) pot varia între populații. În liniile de reproducere, un caz comun este consangvinizarea, care epuizează varianța genetică: ca rezultat, o trăsătură poate avea aceeași medie în două populații, dar are o variație genetică diferită. Abilitatea unui crescător de a selecta o trăsătură este, prin urmare, o caracteristică a liniei, nu a trăsăturii.,
relația dintre părinți și copii este măsurată prin heritabilitate h2. Acesta este definit ca raportul dintre varianța genetică aditivă a dispersiei totale :
h2 = Vga/Vt

(2) corelația 0, unde mărimea copiilor este independentă de cea a părinților. Un părinte mediu valoros a produs un copil de valoare indiferentă, în distribuția copiilor. Coordonatele punctelor sunt situate pe un cerc; așteptarea valorii lor este situată pe o linie paralelă cu axa părinților (d)., (3) corelația părinte-copil intermediar între 0 și 1: coordonatele punctelor sunt situate pe o elipsă. Așteptarea valorii lor nu mai este localizată pe bisectorul axelor de coordonate (care este și axa majoră a elipsei de distribuție), ci pe linia de regresie r. un părinte mediu al valorii a produs un copil cu valoarea b (între 0 și a) cu o anumită variație. Se observă că cazul (3) este intermediar între cazurile (1) și (2)
este prezentat sub diferite ipoteze din Figura 5., Heritabilitatea este capacitatea de a determina dacă o caracteristică poate fi selectată. Figura 5-3 arată că un crescator va fi capabil, într-o trecere generație, pentru a muta medie de un personaj de la valoarea 0 la valoarea b dacă se selectează persoane fizice cu o valoare. Putem arăta că b = h2a. Pentru a selecta o trăsătură, este util pentru a crește heritabilitate raport prin standardizarea mediului (reducând astfel impactul asupra mediului de varianță), și prin plasarea de sine în condițiile de mediu care favorizează apariția trăsătură., Geneticianul Waddington a arătat că, în natură, o schimbare a mediului poate dezvălui astfel noi trăsături care vor fi apoi selectate în timpul evoluției.
referințe și note
Imaginea de copertă. Cochilii de melci Cepaea nemoralis & Cepaea hortensis. André Künzelmann, UFZ.,
Această formulă poate fi scrisă într-un mod mai convențional ca: \pi =\frac{n(n-1))}{2L}\sum _{i=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
Fiind un eșantion de indivizi în care caracterul studiat a fost măsurată, variația poate fi definită ca diferența între valoarea medie de piata de valori și piața de valori. Această măsură, care este întotdeauna pozitivă, indică dispersia indivizilor.aceasta este heritabilitatea în sensul strict al termenului. Heritabilitatea într-un sens larg este raportul dintre suma varianțelor genetice și varianța totală., Această formulă oferă heritabilitate cu ambii părinți („părintele mediu”); estimarea cu un singur părinte ar da h2/2.
Mediu Enciclopedia de Mediu de către Asociația des Encyclopédies de l ‘Environnement et de l’ Énergie (www.a3e.fr), legată la Universitatea din Grenoble Alpes și Grenoble INP, și sponsorizat de Academia franceză de Științe.pentru a cita acest articol: VEUILLE Michel (2021), polimorfism Genetic și variație, enciclopedia mediului, url : https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
articole în Enciclopedia de Mediu sunt puse la dispoziție în conformitate cu termenii licenței Creative Commons BY-NC-SA de licență, care autorizează reproducerea obiectul: citând sursa, nu face uz comercial dintre ei, schimbul de condiții inițiale identice, care reproduce la fiecare reutilizare sau distribuție auzul acestui Creative Commons BY-NC-SA de licență.
Lasă un răspuns