DNA och RNA
de två huvudtyperna av nukleinsyror är deoxiribonukleinsyra (DNA) och ribonukleinsyra (RNA). DNA är det genetiska materialet som finns i alla levande organismer, allt från encelliga bakterier till multicellulära däggdjur. DNA finns i kärnan av eukaryoter och i organeller, kloroplaster och mitokondrier. I prokaryoter är DNA inte inneslutet i ett membranöst kuvert.
hela det genetiska innehållet i en cell är känt som dess genom, och studien av Genom är genomik., I eukaryota celler men inte i prokaryoter bildar DNA ett komplex med histonproteiner för att bilda kromatin, substansen i eukaryota kromosomer. En kromosom kan innehålla tiotusentals gener. Många gener innehåller information för att göra proteinprodukter; andra gener kod för RNA-produkter. DNA kontrollerar alla cellulära aktiviteter genom att vrida generna ” på ”eller” av.”
den andra typen av nukleinsyra, RNA, är mestadels involverad i proteinsyntes. DNA-molekylerna lämnar aldrig kärnan utan använder istället en mellanhand för att kommunicera med resten av cellen., Denna mellanhand är messenger RNA (mRNA). Andra typer av RNA—liknande rRNA, tRNA och microRNA—är involverade i proteinsyntes och dess reglering.
DNA och RNA består av monomerer som kallas nukleotider. Nukleotiderna kombineras med varandra för att bilda en polynukleotid, DNA eller RNA. Varje nukleotid består av tre komponenter: en kvävebas, en pentos (fem-kol) socker och en fosfatgrupp (figur \(\PageIndex{1}\)). Varje kvävebas i en nukleotid är fäst vid en sockermolekyl, som är fäst vid en eller flera fosfatgrupper.,
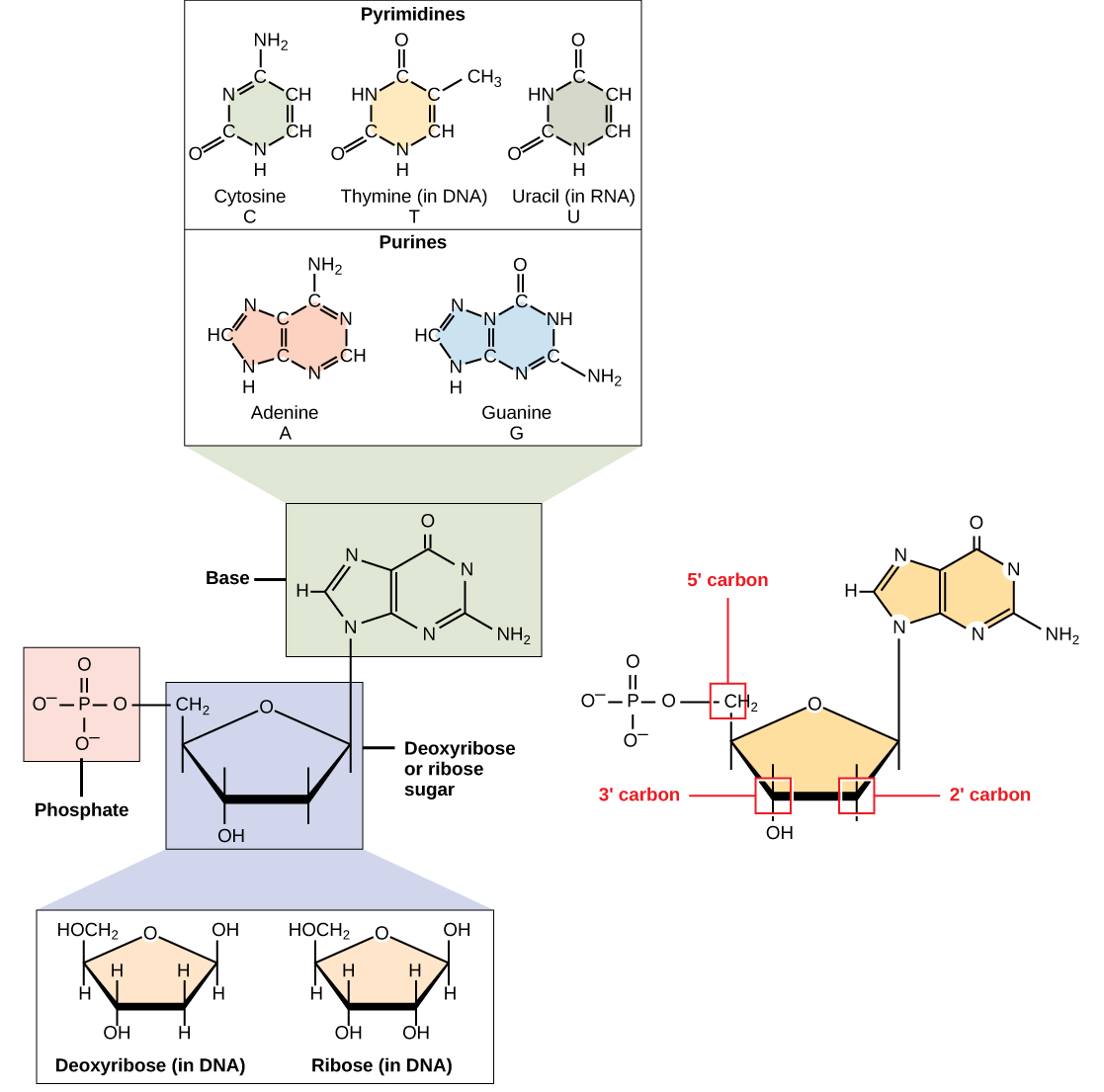
kvävebaserna, viktiga komponenter i nukleotider, är organiska molekyler och är så namngivna eftersom de innehåller kol och kväve. De är baser eftersom de innehåller en aminogrupp som har potential att binda ett extra väte, och därmed minskar vätejonkoncentrationen i sin miljö, vilket gör den mer grundläggande. Varje nukleotid i DNA innehåller en av fyra möjliga kvävebaser: adenin (a), guanin (G) cytosin (C) och tymin (T).
adenin och guanin klassificeras som puriner., Den primära strukturen hos en purin är två kolkväveringar. Cytosin, tymin och uracil klassificeras som pyrimidiner som har en enda kolkvävering som sin primära struktur (figur \(\PageIndex{1}\)). Var och en av dessa grundläggande kolkväveringar har olika funktionella grupper kopplade till den. I molekylärbiologi stenografi är kvävebaserna helt enkelt kända av deras symboler A, T, G, C och U. DNA innehåller A, T, G och C medan RNA innehåller A, U, G och C.
pentossockret i DNA är deoxiribos och i RNA är sockret ribos (figur \(\PageIndex{1}\))., Skillnaden mellan sockerarterna är närvaron av hydroxylgruppen på det andra kolet av ribos och väte på det andra kolet av deoxiribos (så deoxiribos saknas ”an-OH-gruppen). Kolatomerna i sockermolekylen är numrerade som 1′, 2′, 3′, 4′, och 5 ’(1’ läses som ”en prime”). Fosfatrester är fäst vid hydroxylgruppen i 5 ’kolet av ett socker och hydroxylgruppen i 3′ kolet av sockret i nästa nukleotid, som bildar en 5’ -3 ’ fosfodiesterbindning., Fosfodiesterlänkningen bildas inte genom enkel dehydreringsreaktion som de andra kopplingarna som förbinder monomerer i makromolekyler: dess bildning innebär avlägsnande av två fosfatgrupper. En polynukleotid kan ha tusentals sådana fosfodiesterlänkar.
Chargaffs regler
När Watson och Crick fastställde på 1940-talet för att bestämma DNA-strukturen var det redan känt att DNA består av en serie fyra olika typer av molekyler, kallade baser eller nukleotider: adenin (a), cytosin (C), tymin (T), guanin (G)., Watson och Crick kände också till Chargaffs regler, som var en uppsättning observationer om den relativa mängden av varje nukleotid som var närvarande i nästan alla DNA-extrakt. Chargaff hade observerat att för varje given art var överflöd av A densamma som T, och G var densamma som C. Detta var viktigt för Watson & Cricks modell.
exempel \(\Pageindex{1}\)
Chargaff bestämde sammansättningen av nukleinsyror i prover från en mängd olika arter, inklusive prokaryoter och eukaryoter. I ett bakterieprov var andelen adenin 15.,5 %( uppgifter anpassade från Vischer m. fl., 1949). Vilken andel av guanin skulle ha varit närvarande i detta prov och varför?
lösning
eftersom ett par med T, bör mängden T vara ungefär lika med A, eller ungefär 15,5% procent. Således A + T = 15,5 + 15,5 = 31%.
procent av G + C = 100% – 31% = 69%. Eftersom g par med C, bör mängden av var och en av dessa vara ungefär lika, så ungefär 34,5% vardera.,
DNA-Dubbelhelixstruktur
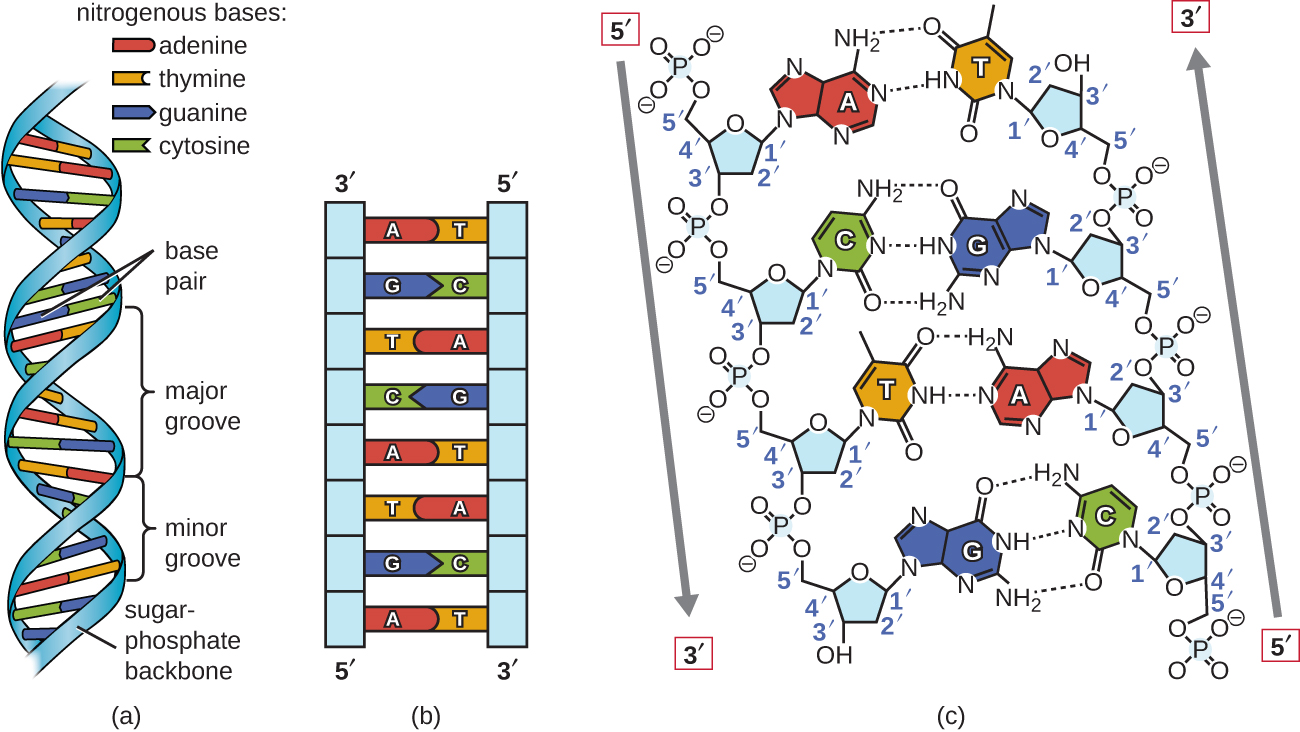
med hjälp av proportionella metallmodeller av de enskilda nukleotiderna härledde Watson och Crick en struktur för DNA som överensstämde med Chargaffs Regler och med röntgenkristallografidata som erhölls (med viss kontrovers) från en annan forskare som heter Rosalind Franklin. I Watson och Cricks berömda double helix innehåller var och en av de två strängarna DNA-baser kopplade genom kovalenta bindningar till en sockerfosfatbackben., Eftersom en sida av varje sockermolekyl alltid är ansluten till den motsatta sidan av nästa sockermolekyl, varje sträng av DNA har polaritet: dessa kallas 5 ’(5-prime) änden och 3 ’ (3-prime) änden, i enlighet med nomenklaturen av kol i sockerarterna. De två delarna av den dubbla spiralen löper i antiparallella (dvs. motsatta) riktningar, med 5′ änden av en sträng intill 3’ änden av den andra strängen. Den dubbla helixen har en högerhänt vridning (snarare än den vänstra vridningen som ofta representeras felaktigt i populära medier)., DNA-baserna sträcker sig från ryggraden mot mitten av helixen, med ett par baser från varje sträng som bildar vätebindningar som hjälper till att hålla de två strängarna ihop. Under de flesta förhållanden är de två strängarna något förskjutna, vilket skapar ett stort spår på ena sidan av dubbelspiralen och ett mindre spår på den andra. På grund av basernas struktur kan A endast bilda vätebindningar med T, och G kan bara bilda vätebindningar med C (kom ihåg Chargaffs regler)., Varje sträng sägs därför vara ett komplement till den andra, och så varje sträng innehåller också tillräckligt med information för att fungera som en mall för syntes av den andra. Denna kompletterande Redundans är viktig i DNA-replikering och reparation.
endast vissa typer av basparning är tillåtna. Till exempel kan en viss purin bara para med en viss pyrimidin. Detta innebär att en kan para med T, och G kan para med C, som visas i Figur \(\PageIndex{2}\). Detta kallas den grundläggande kompletterande regeln. Med andra ord kompletterar DNA-strängarna varandra., Om sekvensen av en sträng är AATTGGCC, skulle den kompletterande strängen ha sekvensen TTAACCGG. Under DNA-replikering kopieras varje sträng, vilket resulterar i en dotter DNA dubbel helix innehållande en föräldra DNA-sträng och en nyligen syntetiserad sträng.
tänka framåt övning \(\PageIndex{1}\)
en mutation inträffar, och cytosin ersätts med adenin. Vilken inverkan tror du att detta kommer att ha på DNA-strukturen?,
svar
adenin är större än cytosin och kommer inte att kunna basera par ordentligt med guaninen på den motsatta strängen. Detta kommer att få DNA att bukta. DNA-reparationsenzymer kan känna igen utbuktningen och ersätta den felaktiga nukleotiden.
Lämna ett svar