
genetisk polymorfism är förekomsten av alternativa tillstånd av DNA, bestämma variationen av organismens högre nivåer av integration. Det finns olika typer av genommodifieringar (mutationer). De mest studerade är nukleotid substitutioner i kodning och regulatoriska regioner.
Definition
genetisk polymorfism är förekomsten, i en population, av flera alternativa tillstånd av DNA, eller alleler, vid en definierad position i genomet, eller lokusposition av genen på kromosomen., I populationsgenetik, en uppsättning homologa gener (homologiklass). Två gener är homologa om de matchar meios. Loci.. Denna definition har flera aspekter:
(1) för det första måste egenskapen bäras av kromosomerna och vara överförbar.
(2) då måste allelerna vara homologatvå kromosomer eller två gener sägs vara homologa om de matchar och ömsesidigt utesluter varandra från meios. för deras position i genomet, vilket utesluter vissa typer av variation som kommer att diskuteras senare.,
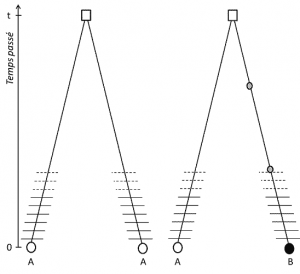
(3) men eftersom egenskapen är överförbar innebär positionshomologi också att allelerna är homologa genom nedstigning.om de är olika är det därför att en mutation (åtminstone) har inträffat i en av de linjer som länkar dem till sin sista gemensamma förfader (Figur 1).
(4) Slutligen kan genetisk polymorfism definieras på skalan av den minsta enhet som komponerar DNA: nukleotidplatsen., Som ett resultat kan varje nukleotidvariant bestämma polymorfismen hos de högre nivåerna av biologisk struktur – individens gen, protein och fenotyp-som blir skalor där genetisk polymorfism kan beskrivas (Figur 2).
ta exemplet med ABO – blodsystemet, vilket motsvarar antigenmotivdel av antigenet som erkänns av en antikropp eller lymfocytreceptor. Även känd som en epitop eller antigendeterminant. Samma antigen kan ha flera epitoper (identiska eller olika) och därmed inducera ett varierat immunsvar., involverad i kompatibiliteten under blodtransfusioner. Detta är ett fall av genetisk polymorfism som finns i mänskliga populationer, som involverar proteinet ABO glykosyltransferaseenzym som möjliggör överföring av rester som innehåller sockerarter till proteiner. När det gäller ABO-systemet inducerar glykosyltransferas a respektive B en individ att tillhöra grupp A eller B. När båda glykosyltransferaserna är närvarande är individen av grupp AB. som har tre alleler, A, B och O. en individ kan ha (AA), (AO); (BB), (BO); (OO) eller (AB) genotyper., Dess fenotyp kommer då att vara,,;,; eller, där vi ser att A och B ”dominerar” O (dvs deras uttryck masker som av O), och att de är co-dominerande sinsemellan (den heterozygot mellan A och B har ett igenkännligt uttryck,).
en polymorfism kan beskrivas först vid skalan av DNA-sekvensen hos locus som kodar för ett protein. Vissa polymorfismer är ”synonyma”, d.v. s. de ändrar inte aminosyrasekvensen av proteinet; de är ofta de mest talrika polymorfismer. De andra modifierar aminosyror och kallas ”ersättning” polymorfismer., Substitutionspolymorfismer av ABO glykosyltransferas är av två typer: polymorfismer som förändrar aminosyran men inte den antigena enheten (de stör inte abo-fenotypen hos individer); och andra ersättningspolymorfismer, som bestämmer ABO-fenotypuppsättningen av observerbara egenskaper hos en individ..
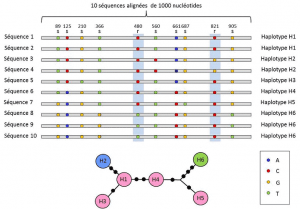
mellan nukleotid-och allelnivåerna tar genetiska analyser hänsyn till en mellanliggande nivå av Beskrivning: det linjära arrangemanget av variabla platser på platsen. Dessa arrangemang, kallade haplotypergrupp av alleler av olika loci som ligger på samma kromosom och överförs vanligtvis tillsammans. Haplotype är en bildad av sammandragningen av den engelska frasen haploid genotyp eller haploid genotyp., Alla gener som finns på samma kromosom och vars alleler segregerar tillsammans under meios utgör en haplotype. Dessa gener kallas ”genetiskt kopplade”, är användbara i evolutionär genetik eftersom de tillåter att hitta de genealogiska länkarna mellan allelerna (Figur 2).
Nukleotidpolymorfismer finns också i intergena regioner (utanför kodningsområden). Vissa påverkar de områden där genuttryck regleras och därför har fenotypiska uttryck. Andra har ingen känd effekt och kallas ”tyst”.,
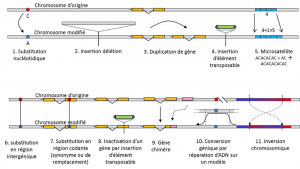
förutom nukleotidsubstitutioner hänvisar polymorfism för en viss typ av mutation, mikrosatelliter, till variationer i längden av ett ofta icke-kodande DNA-fragment (Figur 3) på grund av en förändring av antalet repetitioner i en kort upprepad sekvens av nukleotider, t.ex. CACACACA eller TGTGTG.
inte all genetisk variation faller inom definitionen av genetisk polymorfism, eftersom det kräver positionshomologi av alternativ (Figur 3)., Således upprepas sekvenser erhållna genom duplicering av en gen, för vilken två repetitioner inte är homologa per position. Ofta kan vi inte heller prata om homologi för överförbara element, eftersom de i allmänhet ändrar position under cellgenerationer, kan multiplicera och invadera genomet. Homologin hos två överförbara element kan inte definieras. Å andra sidan motsvarar två kodningssekvenser av samma lokus, varav den ena är intakt och den andra inaktiverad genom införande av ett transposerbart element, väl en homologisituation., En dag kommer vi förmodligen att överväga att prata om polymorfism för modifiering av så kallade ”epigenetiska” kromosomer, som ibland överförs över flera cellgenerationer, somatisktkvalificerar icke-reproduktiva celler eller soma. En somatisk mutation, som påverkar en gen av en somatisk cell, försvinner med bärarindividen. eller germinalkvalificerar gameter. En germlinmutation kan överföras till dess avkomma. såsom metylering. Deras betydelse för utvecklingen av populationer återstår att bedöma., Begränsningen av begreppet genetisk polymorfism till vissa kategorier av variation-i huvudsak nukleotid substitutioner-kommer från det stora nyttan av den senare att utforska historien om naturliga populationer. De lånar sig till matematisk modellering av evolutionen.
ordet polymorfism betyder ”flera former”. Det motsätter sig monomorfism, vilket indikerar frånvaron av variation. I biologins ordförråd har denna opposition länge använts, före genetiken, för att signalera samexistensen av flera typer av individer i samma art, t. ex., caste polymorfism (drottning och arbetare) i sociala insekter; säsongspolymorfism (pälsförändringar) hos vissa arktiska däggdjur. Dessa fall faller inte under genetisk polymorfism, ett nyare koncept med en smalare och mer exakt mening. Kontinuerliga variationer (t.ex. storleksvariationer) ingår inte heller i kategorin polymorfism, eftersom de inte innehåller några tydliga alternativ. Men locus som påverkar storleken faller inom denna definition., Vi kommer att diskutera nedan de mest studerade genetiska variationerna eftersom de är de viktigaste för evolution, nukleotidsubstitutioner, och då kommer vi att diskutera variationen av fenotyper.
mätning
det räcker inte att säga att ett lokus är mer eller mindre variabelt utan att ge en kvantitativ uppskattning till denna dom. En forskare kan studera variationen på olika skalor. Om han bara är intresserad av alleler av ett protein, kommer han att mäta den alleliska mångfalden, symboliserad av ”H”. Om han är intresserad av DNA-mångfald, kommer han att mäta nukleotiddiversitet, symboliserad av ”π”.,
allelens mångfald h definieras som sannolikheten att dra två olika alleler under två provtagningar med ersättningutföra en successiv dragning med leverans av p-tokens i en urn som innehåller N-tokens, innebär att du tar en första token, läser dess värde, lägger den tillbaka i urnan, tar en andra token, läser dess värde, lägger den tillbaka i urnan etc. tills pth-token. Det innebär att välja p-objekt bland n med repetition (du kan välja samma objekt flera gånger) och i ordning (den ordning du väljer objekten är viktig)., Antalet successiva dragningar med tokens bland n är: n × n × n × … × n = NP.. Om vi kallar pi frekvensen av allelen av rank i, visar vi att sannolikheten att plocka två gånger samma allel är F = Σ pi2. Provets alleliska mångfald är då dess komplement till 1, dvs.:
H = 1 – Σ pi2 (1)
detta gäller både proteinalleler och haplotyper. Detta kallas haplotypisk mångfald. Det kan också kallas heterozygositet, eftersom det i fallet med en diploid locus ger den förväntade frekvensen av heterozygoter.,
nukleotiddiversiteten π motsvarar h på skalan för varje nukleotid. Det beräknas som genomsnittet av antalet nukleotidskillnader mellan sekvenser i ett prov som tagits två av två (δij), dividerat med längden av DNA-fragmentet i antalet nukleotider (L).
π = medelvärde (δij) /L (2) (Se referens)
detta värde skiljer sig åt mellan arter. I de kodande delarna av det mänskliga genomet skiljer sig två kromosomer slumpmässigt utvalda från befolkningen i genomsnitt med en nukleotid per tusen., I fruktflugan (Drosophila melanogaster) är denna skillnad ungefär en av hundra. Flugan är alltså tio gånger mer variabel än människan. När dessa värden är relaterade till kodningsregionernas storlek (cirka 15 500 gener i Drosophila, minst 22 000 hos människor), eller till och med genom (140 miljoner baspar per haploidgenom i flugor, cirka 3 200 miljoner eller tjugo gånger mer hos människor), är antalet polymorfa platser astronomiska och resulterar i att ingen föds av den sexualiserade generationen i någon art är, har varit och kommer att vara genetiskt identisk med en annan tidigare, nuvarande eller framtida., Informationskraften hos DNA-polymorfism är enorm. I rättsmedicin kan detektiver identifiera alla misstänkta från sexton mikrosatellit locuses.
historia
termen variation är i titeln på de två första kapitlen i Darwins bok The Origin of Species (1859). Darwin introducerade denna uppfattning i naturvetenskapen vid en tidpunkt då lagarna om biologisk ärftlighet förblev ett mysterium och ägnade en annan viktig bok till den, Variation i djur och växter (1868)., Övertygad om att evolutionen handlade om små variationer som gjorde liten skillnad för organismernas anpassning till deras levnadsförhållanden, fäste han stor vikt vid små kvantitativa variationer, vilket fick hans efterträdare (särskilt Karl Pearson) att hitta biometryscience of life measurement. Avser i mycket bred mening den kvantitativa studien av levande varelser.. Men återupptäckandet av Mendels lagar 1900 skiftade intresset till diskontinuerliga variationer.
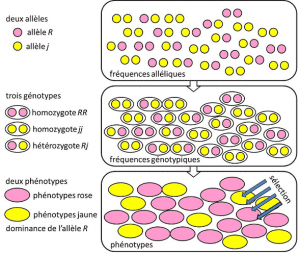
från 1908 till 1930 var populationsgenetik i huvudsak en teoretisk disciplin som försökte förena Darwinism med mendelisk ärftlighet baserad på överföring av en enda gen under ett dominerande, recessivt eller könskromosombundet läge X (eller Y). Avser ett genetiskt drag med enkel determinism, av ett par eller ett litet antal par av gener., och där sannolikheter spelade en viktig roll. Genetik är kontraintutiv., Det förutspår att det inte finns någon reproduktion av föräldern i hans efterkommande, eftersom den senare genotypen härrör från lotteriet om separationen av allelerna före sammansmältningen av de två föräldrahalvgenomerna. Det var insett att dessa är de alleliska frekvensernafrekvens där allelen av en variant finns i en befolkning. Uttryckt som andel eller procentandel. Summan av allelfrekvenserna för alla alleler av en gen i en population är därför per definition lika med 1. I populationsgenetik representerar allelfrekvenser genetisk mångfald på populations-eller artsnivå., som överförs mellan generationer, inte genotyper eller fenotyper. Dessa frekvenser är mer eller mindre stabila från en generation till nästa och producerar, utöver den multipla sammanflätningen av filiationer mellan individer, samma genotypiska frekvensergenetisk struktur hos befolkningen. Bestämd från allelfrekvenserna (Figur 4). Fördelningen av populationsgenotyper, associerad med varians, är därför den enda förutsägbara faktorn., Omkring 1930 bidrog de tre teoretikerna Ronald Fisher, JBS Haldane och Sewall Wright till att skapa begreppet mendelisk befolkningbefolkningbefolkning vars ärftlighet lydde Mendels lagar. . I detta sammanhang associerar evolutionen nödvändigtvis tre strukturella nivåer: genen, individen och befolkningen. Denna trevägssamband kan sammanfattas med följande formel: populationsutveckling är en förändring av allelfrekvensen (genskala, men också gruppskala), en förändring som beror på sortering av fenotyper genom urval (individuell skala)., Empiriska populationsgenetik forskning för att sedan utvecklas, men var begränsad under en lång tid av vår okunskap om hur kromosomerna arbete (DNA-strukturen var förstås 1953 och dess sekvensering började 1977), och var tvungen att falla tillbaka på studier av några synliga polymorfismer, såsom färgning av nyckelpigas elytra eller skalet av sniglar (se Fokus Den stora snigeln debatt). Det fanns mycket debatt bland genetiker om huruvida naturliga populationer i allmänhet var polymorfa eller monomorfa, och om polymorfism i sig var fördelaktig eller inte., Studien av makromolekylär skalvariation fick vänta till 1966 (för proteiner) och 1983 (för DNA) innan de besvarade dessa frågor.
kvantitativa variationer och heritability
När vi tittar omkring oss ser vi fysiska skillnader mellan varelser, några komplexa som ansiktsdrag, andra lätta att mäta som vikt eller höjd. Sunt förnuft tyder på att de delvis är överförbara, men på ett sätt som är svårt att specificera. Dessa egenskaper är ofta polygeniskaunder beroendet av många gener. Vi pratar om polygenisk ärftlighet. Diabetes är en polygenisk sjukdom.,, vilket innebär att de påverkas av många platser. Utvecklings genetik, och dess tillämpning på jämförelsen av arter (evo-devo, för ”utveckling av utveckling”) riva banan av komplexa interaktioner som gör att under utveckling cellerna i kroppen, alla med identiska genetiska bagage, differentiera sig genom Epigenes att ge olika vävnader. Det är fortfarande oklart hur komplexa tecken byggs. Mätbara fenotyper kan studeras med biometri, baserat på egenskaperna hos statistiska fördelningar., Ett värde med användbara egenskaper är varianseni ett prov av individer där ett givet drag mättes är variansen skillnaden mellan medelvärdet av kvadraten av värden och kvadraten av medelvärdet av värdena. Denna åtgärd, som alltid är positiv, indikerar spridningen av individer. . När flera oberoende orsaker bestämmer variationen av en linje, är deras variationer additiva och deras summa ger variansen av linjen. Om de inte är oberoende läggs summan av kovarianserna till summan av varianserna., Den totala variationen Vt av en fenotypisk egenskap är sådan att :
Vt = Vga+Vgd+Vgd+Vgi+Ve
Vga, additiv genetisk varians, är summan av de avvikelser på grund av att varje locus oberoende av andra, Vgd, dominerande genetiska variansen, resultat från samverkan mellan de två alleler i samma locus; Vgi, interaktion variansen mellan locuses, resultaten från det faktum att locuses av samma individ interagerar, Fem, variansen av miljön, är här antas oberoende av genetisk varians.,
den genetiska variansen av dominans är gemensam för två barn av samma föräldrar, eftersom de delar dominans effekter när vid en given locus de har fått samma alleler från båda föräldrarna, vilket händer med dem på en av fyra locus. De delar inte dessa effekter med sina föräldrar, så de är mer ”lika” med varandra än de är för sina föräldrar, trots att de delar så många gener med varandra. Visst kan en förälder av en slump ha fått samma två alleler som ett av sina barn på en given locus., Denna chans beror på frekvensen av alleler i befolkningen. Vi ser att graden av likhet mellan ett barn och hans eller hennes förälder för ett visst drag är en offentlig egendom.
Mer Allmänt kan alla komponenter i genetisk varians (Vga+Vgd+Vgi) variera mellan populationer. I avelslinjer är ett vanligt fall inavel, vilket uttömmer genetisk varians: som ett resultat kan ett drag ha samma genomsnitt i två populationer, men har en annan genetisk variation. En uppfödares förmåga att välja ett drag är därför en egenskap hos linjen, inte av egenskapen.,
förhållandet mellan föräldrar och barn mäts av heritability h2. Det definieras som förhållandet mellan additiv genetisk varians och total varians:
h2 = Vga / vt

(2) korrelation av 0, där storleken på barn är oberoende av föräldrarnas. En värdefull Genomsnittlig förälder har producerat ett barn av likgiltigt värde, inom fördelningen av barn. Koordinaterna för punkterna ligger på en cirkel; förväntan om deras värde ligger på en linje parallellt med föräldrarnas axel (d)., (3) mellanliggande förälder-barn korrelation mellan 0 och 1: koordinaterna för punkterna ligger på en ellips. Förväntan om deras värde ligger inte längre på koordinataxlarnas bisektor (som också är distributionsellipsens huvudaxel), men på regressionslinjen r. en genomsnittlig förälder av värde har producerat ett barn av värde b (mellan 0 och A) med en viss varians. Det bör noteras att fall (3) är mellanliggande mellan fall (1) och (2)
det visas under olika antaganden i Figur 5., Ärftlighet är förmågan att avgöra om en funktion kan väljas. Figur 5-3 visar att en uppfödare kommer att kunna, inom en korsningsgenerering, flytta genomsnittet av ett tecken från värdet 0 till värdet b om han väljer individer med värde a. vi kan visa att b = h2a. för att välja ett drag är det användbart att öka heritabilitetsförhållandet genom att standardisera miljön (vilket minskar miljövariansen) och genom att placera sig i de miljöförhållanden som gynnar egenskapens utseende., Genetiker Waddington har visat att, i naturen, en förändring av miljön kan därmed avslöja nya egenskaper som sedan kommer att väljas under evolutionen.
referenser och anmärkningar
Täckbild. Snigelskal Cepaea nemoralis & cepaea hortensis. André Künzelmann, UFZ.,
denna formel kan skrivas på ett mer konventionellt sätt som: \pi =\frac{n(n-1))}{2l}\sum _{i=1}^{n-1}\sum_{j=i+1}^{n}\delta ij
att vara ett prov av individer i vilka den studerade egenskapen mättes, varians kan definieras som skillnaden mellan medelvärdet av kvadraten av värdena och kvadraten av medelvärdet av värdena. Denna åtgärd, som alltid är positiv, indikerar spridningen av individer.
detta är heritability i strikt bemärkelse. Heritability i bred mening är förhållandet mellan summan av genetiska variationer och total varians., Denna formel ger heritability med båda föräldrarna (Den” genomsnittliga föräldern”); uppskattningen med endast en förälder skulle ge h2/2.
miljö Encyclopedia of the environmental Encyclopedia of the environment by the Association des Encyclopédies de l ’environnement et de l’ énergie (www.a3e.fr), kontraktuellt kopplad till Universitetet i Grenoble Alpes och Grenoble INP, och sponsras av den franska vetenskapsakademin.
för Att citera denna artikel:VEUILLE Michel (2021), Genetisk polymorfism och variation, Uppslagsverk av Miljön, url : https://www.encyclopedie-environnement.org/en/life/genetic-polymorphism-and-variation/.,
artiklarna i miljöens encyklopedi görs tillgängliga enligt villkoren i Creative Commons BY-NC-SA-licensen, som tillåter reproduktion under förutsättning att: citera källan, inte göra kommersiell användning av dem, dela identiska ursprungliga villkor, reproducera vid varje återanvändning eller distribution omnämnandet av denna Creative Commons BY-NC-SA licens.
Lämna ett svar