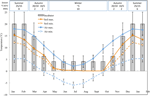
Figur 1. Månatliga uppgifter om högsta och lägsta luft-och marktemperatur i de australiska Alperna (medelvärde ± STDEV). Lufttemperaturer är medelvärden från BOM-databasen, jordtemperaturer mättes med iButtons (se metoder). Barer indikerar experimentella inkubatortemperaturer (medelvärde ± STDEV)., Motsvarande tid på året( säsong), inkubatortemperaturreglering (°C, 12/12 h, ljus/mörk) och behandlingstid (veckor) enligt vad som anges i den övre panelen.
för att upptäcka temperaturförhållanden nära dem som frön upplever efter spridning, insamlades dagliga maximala och minsta jordtemperaturdata inom studieplatsen (Sjömanshut; 36,27°s och 148,17 ° E, 2030 M A. S. L.) med hjälp av iButton dataloggers (n = 10, inbäddade datasystem, USA) placerade 4 cm under markytan vid basen av vegetation från 17 januari till 17 December 2012., Vi placerade loggers på 4 cm för att undvika ytstörningar men fortfarande vara representativa för förhållanden för vilka frön exponeras i jord; detta djup är mellanliggande till vad som har använts i tidigare alpina marktemperaturövervakningsövningar (Scherrer och Körner, 2011; Pauli et al., 2015). Som misstänkt sjönk den genomsnittliga marktemperaturen inte under frysning under vintern (Figur 1). På sommaren är marktemperaturen under vegetation känd för att spåra omgivningstemperaturen, medan temperaturen på bar jord kommer att överstiga omgivningen med upp till 15 ° C (Soil Conservation Service opublicerade register, 1960–1970-talet).,
insamling och groning av utsäde
mogna frön av 54 arter från 16 familjer och 37 släkten samlades mellan januari och April 2009, 2010 och 2011 (se Tabell 1 för fullständiga namn och myndigheter. Kuponger lämnades in på Australian National Herbarium, Canberra). Totalt representerade arten mer än en fjärdedel av den australiensiska angiospermfloran i alpina regioner (Costin et al., 2000, se Tabell 1), även om många av dessa arter sträcker sig till under treeline också., Lönsamheten för alla samlingar uppskattades före sådd i experimentella grobarhetsförhållanden med hjälp av tetrazolium chloride (TZ) staining technique (International Seed Testing Association, 2003). För mer information om insamling och bearbetning, Se bilaga A i kompletterande Material.
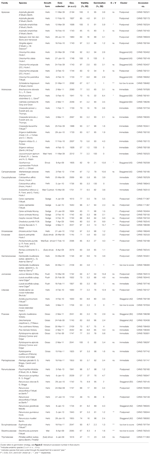
tabell 1. Studieart, familj och myndighet, tillväxtform och samlingsdetaljer, procentuell fröviabilitet (TZ-beräknad, ± S. E.), slutlig procentuell spiring uppnådd (± S. E.,), statistisk skillnad mellan livskraft och grobarhet (g < V, testad endast i fall där grobarheten var mindre än livskraften).
fenologi av frögroning undersöktes genom att härma temperaturregimer som alpina frön upplever på plats, efter spridning, i en artificiellt förkortad progression av årstiderna. Fröer var imbibed under hela experimentet, vilket resulterar i potential för spiring när vatten inte begränsar., Luft-och marktemperaturdata (Figur 1) användes för att styra och validera inkubatortemperaturregimer, även om de logistiska kontrasterna av att arbeta med inkubatorer hindrade införlivande av de fluktuationer som är förknippade med naturliga förhållanden. Grobarhet tester används åtta replikat av 25 frön per samling, sås i 9 cm diameter plast petriskålar innehållande 1% vanligt vatten-agar. Petriskålar förseglades med Parafilm för att undvika agar-uttorkning, innan de placerades i spiringskuvöser (Thermolin Scientific, Melbourne, NSW, Australien)., Hälften av disken var inslagna i aluminiumfolie för att utesluta ljus och ge en indikation på lätta krav på spiring. Var och en av de fyra replikerna per lätt behandling per samling placerades på en annan hylla inom samma inkubator och plats inom den hyllan omarrangerades varje vecka. Alla replikat av varje samling flyttades genom följande simulerade termoperioder: 8 veckor vid 25/15 ° c (dag / natt temperatur) → 2 veckor vid 20/10 ° C → 2 veckor vid 10/5 ° C → 10 veckor vid konstant 4-5 ° C → 2 veckor vid 10/5 ° C → 2 veckor vid 20/10 ° C → 6 veckor vid 25/15 ° C., Således efterliknade en cykel av hela experimentet temperaturer som påminner om sommar, tidig höst, sen höst, vinter, tidig vår, sen vår och sommar och varade totalt 32 veckor. Det förkortade året ansågs rimligt eftersom anslutningarna i allmänhet hade upphört med ytterligare spiring i en given behandling innan de skiftades till nästa ”säsong”., En 12/12 h ljus/mörk fotoperiod tillhandahölls genom fluorescerande rör (> 90 µmol m−2 s−1 i mitten av varje hylla), och under vår -, sommar-och hösttemperaturregimer sammanföll ljuset med dagens varma temperaturperiod. Spiring, definierad som radikelframväxt med mer än 1 mm, gjordes var 7: e dag, och grodda frön och plantor avlägsnades därefter från disken. Spiring av folieförpackade plattor bedömdes som ovan men under mycket lågt grönt ljus.,
efter en experimentell cykel (32 veckor) avslutades de flesta samlingarna (se Kompletterande information för undantag), efter att ha uppvisat nollspiring i minst 2 veckor vid denna tidpunkt. Efter avslutningen dissekerades alla återstående intakta frön med en skalpell under ett mikroskop. Fröer med en fast, färsk frösperm och embryo bedömdes vara livskraftiga och fröer som är tomma från ett embryo drogs av från det totala antalet vid beräkningen av procentuell grobarhet för varje samling., Vi beräknade ett index över de lätta kraven för spiring för varje art som en bråkdel av spiring i den mörka behandlingen vid den tidpunkt då kumulativa spiringsprocentiler på 50 och 95% uppnåddes i den alternerande ljus / mörka behandlingen (se nedan).
ekologiska korrelat av grobarhet strategi
vi sammanställt en databas av arters funktionella egenskaper och geografiska åtgärder från publicerade data för att bedöma korrelationer med grobarhet strategi., Databasen innehöll följande kontinuerliga egenskaper: Genomsnittlig frömassa, lätta krav på spiring (se ovan), specifik bladyta (färskt område/torrmassa), individuell bladyta, individuell bladtorrvikt och höjd vid mognad. De minsta, genomsnittliga och maximala insamlingshöjningarna för arten bestämdes från alla herbariumrekord i det australiensiska virtuella herbariet, och höjden vid vilken de specifika frösamlingarna som användes i studien gjordes inkluderades också som kontinuerliga egenskaper. Tillväxtform gjordes som en av tre kategorier: graminoid, ört eller buske., Vi inkluderade också ett binärt drag: om fröna var endospermiska eller inte.
dragdata samlades in från en mängd olika källor, inklusive Australian Virtual Herbarium (http://avh.chah.org.au/), Australian National Herbarium preparatinformationsregister (https://www.anbg.gov.au/cgi-bin/anhsir), Kosciuszko Alpine Flora (Costin et al., 2000), New South Wales Flora (Harden, 1990), och den jämförande interna morfologin av frön (Martin, 1946). Frömassa och specifikt bladområde erhölls från enskilda fältsamlingar som användes i spridningsförsöken., Frömassa erhölls genom att väga fyra massor av 10 frön. SLA bestämdes baserat på fem fullt utökade blad från separata individer per art. Dessa samlades in i fältet, skannades på en flatbäddsskanner, torkades vid 60 ° C till konstant vikt och vägdes till närmaste 0,000 g. om otillräckligt frö fanns tillgängligt, eller specifik bladyta inte var tillgänglig, erhölls data från andra ANBG-samlingar av samma art eller från andra datakällor (S. Venn, opublicerade data)., Huruvida ett frö var endospermiskt bestämdes från ritningar av fröet i ANGB seed bank records i samråd med anbg seed bank-personal. Den fullständiga databasen och källorna är tillgängliga på begäran från författarna.
dataanalys
Grobarhet mönster utvärderades visuellt och med hjälp av en klusteranalys i PC-ORD för Windows, version 6 (MJM Software Design, Gleneden Beach, eller, USA). Analysen använde en euklidisk avståndsmatris och en närmaste granne som gick med i algoritmen., Datapunkter för klusteranalysen var tiden (veckor) till specifika kumulativa spiringsprocentiler (T20, T40, T50, T60, T80 och t95%) för varje samling (i genomsnitt över de 4 rätterna). Således skulle en samling som grodde tidigt i experimentet ha korta tider till percentiler upp till t80% till exempel, medan en samling som skjutit upp spiring tills temperaturen ökade efter den simulerade vintern skulle ha långa tider till dessa percentiler., Mer komplexa spiringsmönster, såsom långsam, stadig spiring eller sprängningar av spiring, kan också detekteras, och skulle indikeras av jämnt fördelade tider eller tider grupperade i början respektive slutet av experimentet. Denna metod användes som ett sätt att införa en objektiv bedömning av spiringsmönstret, oberoende av temperaturen i sig.,
en envägsanalys av variansen (ANOVA) utfördes för replikatplattorna i varje samling för att bedöma om det fanns en signifikant skillnad mellan procentuell viabilitet (enligt TZ-testet) och slutlig procentuell spiring. Ytterligare allmänna linjära modeller användes för att bedöma om klustren som var resultatet av klusteranalysen skilde sig åt i viabilitet eller slutlig grobarhet.
associationer mellan ekologiska egenskaper och grobarhet strategi kluster testades på två sätt., För det första testade vi för genomsnittliga skillnader mellan kluster med ANOVA med kluster som en fast effekt och ekologisk korrelerar som svarvariablerna, eller med replikerade g-test för endospermy och tillväxtform. För det andra, eftersom spiring och andra funktionella egenskaper är mycket benägna att påverkas av evolutionär historia, bedömde vi dessa korrelationer med hjälp av fylogenetiska regressioner. En fylogeni var konstruerade för studien arter med hjälp Phylocom Phylomatic och bladj packning i R (Webb et al., 2008; R Core Team, 2013)., Vi bedömde närvaron av fylogenetisk struktur i variabler med hjälp av fylosig eller phylo.d (diskreta eller binära variabler) funktion Phytools paketet i R (Revell, 2012). Fylogenetiska regressioner utfördes med pgls-funktionen i Caper (Orme et al., 2013). Analyser utfördes på ett strukturerat sätt som återspeglar klusteringen som avslöjades i PC-Ord (se nedan)., Inledningsvis inkluderade vi fyra spridningsstrategier (uppskjutna, omedelbara och förskjutna kategorier inom var och en av föregående), sedan tre (lumping alla förskjutna arter och jämföra med uppskjutna och mellanliggande) och slutligen bedömde vi skillnader mellan de två bredaste strategierna, uppskjutna och mellanliggande.
resultat
Fröviabilitet jämfört med slutlig Grobarhet
totalt sett var fröviabiliteten och den slutliga grobarheten hög. Den genomsnittliga TZ-beräknade lönsamheten i alla samlingar var 80 ± 1.,7%, med mer än två tredjedelar av samlingarna uppvisar mer än 75% lönsamhet (Tabell 1). Det fanns ingen skillnad i samlingens kvalitet under de tre åren (Genomsnittlig lönsamhet 2009, 2010 och 2011 var 79 ± 2,8, 80 ± 2,7 respektive 81 ± 3,7%), vilket tyder på att bankverksamheten för utsäde som samlades in 2009 inte minskade deras lönsamhet.
Grobarhet som överskred eller inte var signifikant lägre än TZ-beräknad viabilitet uppnåddes för 45 av de 54 arterna (enkelriktad ANOVA: p>0,05, Tabell 1)., Av de återstående nio arterna var den slutliga Groningen av fyra arter mycket låg (< 5%), trots god lönsamhet (ca. 55-94%), och därmed kunde vi inte dra några slutsatser om grobarhet strategier för dessa arter. Underlåtenhet att gro kan återspegla antingen att våra behandlingar inte lindra dvala eller att fröna inte var helt mogna vid insamling. Cut tester i slutet av experimentet visade att i alla fall, de flesta av icke-grodda frön innehöll en frisk, imbibed embryo., Intressant nog uppnådde 34 arter grobarhet som var större än den beräknade TZ-lönsamheten (Tabell 1), vilket tyder på att TZ-testet vanligen underskattade fröviabiliteten.
Grobarhet kluster
klusteranalys avslöjade två stora kluster (A och B), med delstruktur i varje (figur 2). För de flesta arter (n = 21 arter från 10 familjer) sköts Groningen upp till under en period av vintertemperaturer (5°C dag/natt) eller tills temperaturen höjdes till 10/5°C efter vintern (A1 och A1:vinter, Figur 2, 3A)., Alternativt började spiring omedelbart vid 25/15°c (n = 17 arter, sex familjer, sett i kluster B1, figurerna 2, 3B). För de flesta av dessa arter var grobarheten snabb; 50% av den totala grobarheten (t50) inträffade på mindre än 3 veckor (kompletterande Figur 1).
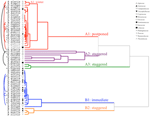
Figur 2. Klusteranalys dendrogram baserat på tid till kumulativa grobarhet percentiler (20, 40, 50, 60, 80, 95%). Arter som anges med släktet initial och första delen av artens namn. Asterisker markerar samlingar som bankades före testning., Fina bågar ansluter dubbla samlingar för att jämföra effekten av Bank på spiring. Symboler betecknar olika familjer (se legend). Samlingar som visas i grå text är de som utsattes för två cykler av experimentet.
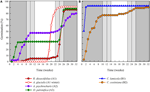
Figur 3. Kumulativ procentuell spiring av en representativ art från varje groddkluster. A) kluster a, arter som skjutit upp grobarheten, b) kluster B, arter som grodde omedelbart., Observera att förskjutna spiringsmönster är kapslade inom ovanstående kluster. Inkubatortemperaturregimer representeras av skuggning (se Figur 1).
inom var och en av dessa breda klasser fanns det dock tydliga delkluster som återspeglade grobarhet som var förskjuten över tiden, dvs. en andel av utsädespartiet grodde före exponering för vintertemperaturer medan resten grodde efter vintern. Förskjuten grobarhet inträffade hos 13 arter från sex familjer och inträffade främst när dagstemperaturen översteg 10 ° C (figurerna 3a och B)., Dessa förskjutna germinatorer delades in i tre grupper i klusteranalysen. Cluster A2 med sju samlingar visade betydande spiring både före och efter den kalla temperaturperioden. Där spiring upphörde före temperaturerna ändras drog vi slutsatsen att bristen på ytterligare spiring före vintern inte drevs av temperatur (A3). Däremot verkade spiring av mycket av A2-gruppen endast stoppas när temperaturen sänktes till 10/5 ° C., Kluster B2 innehöll ytterligare fyra arter som uppvisade betydande grobarhet tidigt i försöket och fortsatte med att uppnå en relativt liten andel av deras totala grobarhet efter att temperaturerna återgått till över 5°C (figur 3b).
jämfört med den relativt små variationen mellan arter inom kluster A1 och B1 visar de förskjutna klustren också mycket större differentiering mellan arter. Individuella kurvor för alla arter visas i Tilläggssiffror 1i-vii. för mer specifika detaljer om grobarhet, se tillägg B i kompletterande Material.,
ekologiska korrelat av grobarhet strategi
av alla ekologiska korrelat som vi ansåg, endast de som är direkt förknippade med fröegenskaper var signifikant korrelerade med grobarhet strategi oavsett om vi bedömde korrelationen i nivå med två stora kluster (skjutas upp mot omedelbar groning), eller i nivå med tre eller fyra kluster, eller om vi stod för fylogenetisk struktur eller inte (Tabell 2)., Arter med uppskjuten spiring hade tyngre frön och var mer benägna att vara endospermiska jämfört med de arter som grodde omedelbart (Figur 4). Arter med förskjuten spiring var mellanliggande i frömassa och endospermy. Men inte alla fröegenskaper var korrelerade med grobarhet strategi: det fanns ingen korrelation mellan lätta krav för frögroning och strategi. I motsats till våra förväntningar var arter med högre specifik bladyta (potentiellt indikativ för högre tillväxttakt) inte mer benägna att uppvisa omedelbar spiring., På samma sätt var arter med högre genomsnittlig höjd eller mindre höjdområden inte mer benägna att uppvisa uppskjuten spiring.

tabell 2. Förekomst av fylogenetisk struktur i, och korrelerar med, grobarhet strategi.
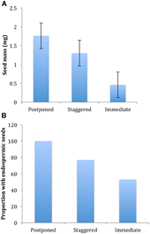
Figur 4., Skillnader i (A) Genomsnittlig frömassa och (B) andel arter med fröplantor bland arter med uppskjuten (kluster A1), förskjuten (kombination av kluster A2, A3 och B2) och strategier för omedelbar grobarhet (kluster B1). Skillnader mellan strategier är signifikanta vid P ≤ 0,05, se Tabell 2.
diskussion
grobarhet i alpina livsmiljöer ansågs historiskt vara miljökontrollerad, med vinter snö isolerande frön mot potentiella groning ledtrådar och gör dvala onödigt (Billings och Mooney, 1968)., Våra resultat stöder dock nyare bevis på att vilande mekanismer är vanliga bland alpina arter både i Australien (Hoyle et al., 2013; Sommerville et al., 2013) och på andra håll runt om i världen (Cavieres och Arroyo, 2000; Schutz, 2002; Shimono och Kudo, 2005; Mondoni et al., 2009; Schwienbacher et al., 2011)., Dormancy verkar spela en viktig roll för att kontrollera in situ-tidpunkten för spiring av många Australiensiska alpina arter, som verkar för att fördröja, skjuta upp eller sakta ner spridningshastigheten och därmed potentiellt ge risk-inversa regenereringsstrategier i en hård och variabel miljö. Kanske mest slående av våra spridningsresultat var andelen arter för vilka spridningsstrategi varierade inom en frösamling för en viss art vilket tyder på att både vilande och icke-vilande egenskaper uppvisades inom samma frösamling av dessa arter., I synnerhet var spridningsstrategier i allmänhet högt bevarade inom familjer och korrelerade också med element av fröanatomi: massa och endospermy. Grobarhet strategier var dock oberoende av andra ekologiskt signifikanta funktionella egenskaper hos den mogna växten.
variation i Grobarhet strategi
frön av nästan hälften av de studerade arter verkade oförmögna att gro tills de utsattes för en sval, våt period (konstant 5°C), vilket tyder på att kall stratifiering lindrade en fysiologisk dvala mekanism., Att skjuta upp spiring till nästa vår kan göra det möjligt för plantor att undvika att etablera sig över eller före den hårda vintern, samtidigt som man optimerar den korta kommande växtsäsongen. Däremot kan den opportunistiska spiring som uppvisas av de arter som groddar omedelbart ge en selektiv fördel när risken för vinterplantordödlighet är låg, genom att växter kan blomma tidigare följande vår eller i större storlek (Donohue, 2002).,
hittills har det inte förekommit några publicerade bevis på intraspecifik variation i vilande mekanismer inom alpina frösamlingar, vilket skulle förklara de förskjutna grobarhet strategier som observerats här. Groning förskjuten över tiden kan också förklaras av varierande nivåer av utsäde mognad bland individer i befolkningen vid tidpunkten för insamling, och / eller kan återspegla en dimorfisk strategi inom en enda anläggning i samband med position på anläggningen eller tidpunkten för utveckling.,
evolutionär bet-hedging framkallas ofta för att förklara den diversifierade strategin i fröspiringsegenskaper (Simons, 2011), såsom den som demonstreras av samlingarna med förskjuten spiring. Om en diversifierad strategi leder till både ökad genomsnittlig lämplighet och en minskning av variansen (t.ex. genom att minska konkurrensen mellan syskon), utgör den dock inte bet-hedging. På samma sätt, om den uppenbara diversifieringen återspeglar skillnader mellan, inte inom, individer är det inte bet-hedging (Starrfelt och Kokko, 2012)., Undersökningar av spridningsmönster för alpina plantor och tolerans för frost-och snösmältning, i kombination med studier av bestämningsfaktorer för spridningsstrategivariation inom arter, kan bidra till att förklara grobarheten hos de arter som uppvisar en förskjuten strategi och kan indikera om de sannolikt kommer att representera bet-hedgingstrategier.,
ekologiska korrelat av grobarhet strategi
vår undersökning av sambandet mellan grobarhet strategi och andra ekologiskt viktiga egenskaper visade att grobarhet drag var korrelerade med andra frö drag men inte med vegetativa egenskaper. Sommerville et al. (2013) fann att Australiensiska alpina örter med icke-endospermiska frön var mer benägna att vara icke-vilande vid spridning. Vårt arbete stöder att hitta och ytterligare indikerar att mindre frön är mer benägna att vara bland de omedelbara germinatörerna. Även Sommerville et al., (2013) poserade inte varför endospermy kan vara associerad med vilande, vi föreslår att placering av reserver i cotyledons kan förbättra tidig tillväxt och etablering i omedelbara groende arter. Våra resultat visade inte någon korrelation mellan lätta krav och viloläge, även om lätta krav varierar mellan arter. Det finns en mängd andra fröegenskaper som buffrar utrotningsrisk i varierande miljöer, till exempel fröspridningsmekanismer och frö livslängd i jordfröbanken (se Tielborger et al., 2012 för en översyn)., I detta skede kan vi inte säga om dessa egenskaper skulle visa korrelationer med grobarhet strategi eller inte.
vi frågade vidare om spridningsstrategin skulle korreleras med ekologiskt viktiga icke-reproduktiva egenskaper, såsom specifikt bladområde eller växthöjd (Westoby, 1998; Wright et al., 2004). I synnerhet vi ansåg att de omedelbara groende arter kan visa drag värden som tyder på snabbare tillväxttakt (med SLA som en proxy) som kan vara nödvändigt för etablering under den korta perioden mellan spridning och uppkomsten av vintern., Men vi hittade inga sådana korrelationer. Huruvida fysiologiska och tillväxtegenskaper hos plantor vid tidiga etableringsstadier kan visa olika mönster än vuxna vegetativa egenskaper är en fråga som är värd att överväga, men bortom det nuvarande arbetet.
slutligen undersökte vi om arter från högre höjder där växtsäsongerna i allmänhet är kortare kan vara mer benägna att uppvisa vilande men det fanns ingen koppling mellan antingen höjdområde, uppsamlingshöjningar eller den genomsnittliga höjden av samlingen för arten som helhet och grobarhet strategi., Tillsammans visar dessa resultat att även om utvecklingen av grobarheten är nära kopplad till ekologiskt viktiga fröegenskaper, innefattar den en till stor del oberoende axel till vegetativa eller fördelningsmässiga egenskaper.
konsekvenser för alpina växtgrupper i ett föränderligt klimat
Australiensiska alpina ekosystem, som alpina områden runt om i världen, hotas av klimatförändringar i kombination med förändringar i brandfrekvens, markanvändningsmönster, tillströmning av invasiva arter och effekterna av ökad mänsklig visitation., Uppvärmningen i samband med klimatförändringen sker snabbare över trädlinjen än vid lägre höjder, och alpina områden förväntas fortsätta att uppleva över genomsnittlig uppvärmning i framtiden (Kullman, 2004). Kapaciteten för fortsatt regenerering via utsäde under nya förhållanden kommer sannolikt att spela en viktig roll när det gäller hur alpina växter reagerar på klimat och därmed sammanhängande förändringar. Dessutom kommer förändrade störningssystem sannolikt att kräva en ökad roll för restaurering och rehabilitering i regionen., De insikter som ges här är därför avgörande för att bevara och förvalta alpina system under förändring. de kan hjälpa till att informera våra förutsägelser om hur studiearter kan påverkas, ligga till grund för planerna för utsädesförökning för dessa arter och användas för att vägleda framtida undersökningar av strategier för grobarhet av alpina frön med identifierbara bevarande-och förvaltningsresultat.,
temperaturförändringar kommer sannolikt att påverka vilande status för frön både före och efter spridning, följaktligen ändra och / eller störa grobarhet strategier och resulterar i förändringar i nuvarande grobarhet fenologi (Hoyle et al., 2008b). När det gäller alpina växter kan klimatuppvärmningen leda till en övergång från vår till höst, främst på grund av förändringar i frödödligheten och leda till stora konsekvenser för arter som för närvarande är anpassade till uppkomst under våren (Mondoni et al., 2012)., Ytterligare forskning om hur förändringar i temperaturer som frön upplever efter spridning, minskad varaktighet av snötäcke och ökad frekvens av extrema temperaturhändelser kommer att påverka spiring krävs för att förutsäga hur arter med viloläge kommer att reagera på klimatförändringarna. För de alpina arter som gror på hösten, före frost och snöfall, tar våra resultat upp frågan om hur plantor av dessa arter kommer att klara av den förväntade minskningen av nederbörd och ökad frekvens av A-säsongens frosthändelser.,
slutligen väcker våra resultat viktiga frågor om variation inom arter, särskilt i samband med att förutsäga klimatförändringens inverkan eller välja vilka frön som ska användas för restaurering. Grobarhet strategier kan variera inom arter beroende på population, blomningstid, frömassa, föräldrainvesteringar, klimat, höjd och/eller mödramiljö och denna variation kan återspegla genetiska, epigenetiska och/eller miljöfaktorer (Cochrane et al., 2015)., Öka höjd står för variation i germinability och/eller dvala status för flera alpina arter (Billings och Mooney, 1968; Dorne, 1981; Cavieres och Arroyo, 2000), men se (Hoyle et al., 2014). Minskad vilande status kan bero på tidig uppsägning av fröutveckling till följd av miljöbelastningar (Steadman et al., 2006; Hoyle et al., 2008b)., Undersökning av variationen i spridningsstrategier inom arter kan avslöja möjligheten för arter att hantera och anpassa sig till förändringar i klimatet, vilket spelar en viktig roll för att bestämma framtida överlevnad och artfördelningar och ge viktig insikt för förvaltning och bevarande.
slutsatser
vårt tillvägagångssätt har avslöjat utbredd förekomst av viloläge i den australiensiska alpina floran och visar att spridningsstrategier varierar inom en frösamling, liksom mellan samlingar., Vidare har vi visat att medan dormancy strategi är fylogenetiskt bevarad och korrelerad med frömassa och endospermy, är det till stor del oberoende av vegetativa egenskaper och range egenskaper. Även om man inte kan extrapolera direkt från våra resultat hur klimat och tillhörande förändringar kommer att påverka alpina floran, kan dessa resultat ligga till grund för utformningen av förökning planer och ytterligare experiment på effekterna av förändrade klimatförhållanden., I synnerhet förespråkar vi ytterligare forskning om att förstå orsakerna till variation inom arter i grobarhet strategi och den roll detta kan spela i förmågan hos arter att motstå förutsagda klimatförändringar.
datatillgänglighet
fullständig grobarhet data och drag databas kommer att lämnas in på DRYAD och Atlas of Living Australien.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.,
bekräftelser
Vi erkänner finansiering från Australian Research Council (LP0991593) och Australian National Botanic Garden och vänner därav. Vi tackar J. McAuliffe, S. Fethers, och S. Lawatsch av ANBG. Vi tackar också M. Holloway-Phillips, M. Cardillo och S. Cunningham för input på tidigare versioner av manuskriptet och för analytisk rådgivning och S. Venn för tillgång till dragdata.
kompletterande Material
Foley, M. E. (1992). Effekten av lösliga sockerarter och gibberellinsyra vid brytning av embryon från utskurna vilda havre (Avena fatua)., Ogräs Sci. 40, 208–214.
Google Scholar
Harden, G. J. (1990). Flora av New South Wales. Sydney, NSW: Royal Botanic Gardens.
Google Scholar
International Seed Testing Association. (2003). Arbetsblad om Tetrazoliumtest. Ista.
Kullman, L. (2004). Alpvärldens föränderliga ansikte. Glob. Byt Nyhetsbrev. 57, 12–14.
Google Scholar
Lämna ett svar