DNA und RNA
Die beiden Haupttypen von Nukleinsäuren sind Desoxyribonukleinsäure (DNA) und Ribonukleinsäure (RNA). DNA ist das genetische Material, das in allen lebenden Organismen vorkommt und von einzelligen Bakterien bis zu mehrzelligen Säugetieren reicht. DNA findet sich im Kern von Eukaryoten und in den Organellen, Chloroplasten und Mitochondrien. Bei Prokaryoten ist die DNA nicht in einer membranösen Hülle eingeschlossen.
Der gesamte genetische Inhalt einer Zelle ist als Genom bekannt, und das Studium von Genomen ist Genomik., In eukaryotischen Zellen, aber nicht in Prokaryoten bildet DNA einen Komplex mit Histonproteinen, um Chromatin zu bilden, die Substanz von eukaryotischen Chromosomen. Ein Chromosom kann Zehntausende von Genen enthalten. Viele Gene enthalten die Informationen, um Proteinprodukte herzustellen; andere Gene kodieren für RNA-Produkte. DNA steuert alle zellulären Aktivitäten, indem Sie die Gene „ein“ oder „aus“.“
Die andere Art von Nukleinsäure, RNA, ist hauptsächlich an der Proteinsynthese beteiligt. Die DNA-Moleküle verlassen niemals den Kern, sondern verwenden stattdessen einen Vermittler, um mit dem Rest der Zelle zu kommunizieren., Dieser Vermittler ist die messenger-RNA (mRNA). Andere Arten von RNA-wie rRNA, tRNA und microRNA—sind an der Proteinsynthese und ihrer Regulation beteiligt.
DNA und RNA bestehen aus Monomeren, die als Nukleotide bekannt sind. Die Nukleotide verbinden sich zu einem Polynukleotid, DNA oder RNA. Jedes Nukleotid besteht aus drei Komponenten: einer stickstoffhaltigen Base, einem Pentose-Zucker (fünf Kohlenstoff) und einer Phosphatgruppe (Abbildung \(\pageIndex{1}\)). Jede stickstoffhaltige Base in einem Nukleotid ist an ein Zuckermolekül gebunden, das an eine oder mehrere Phosphatgruppen gebunden ist.,
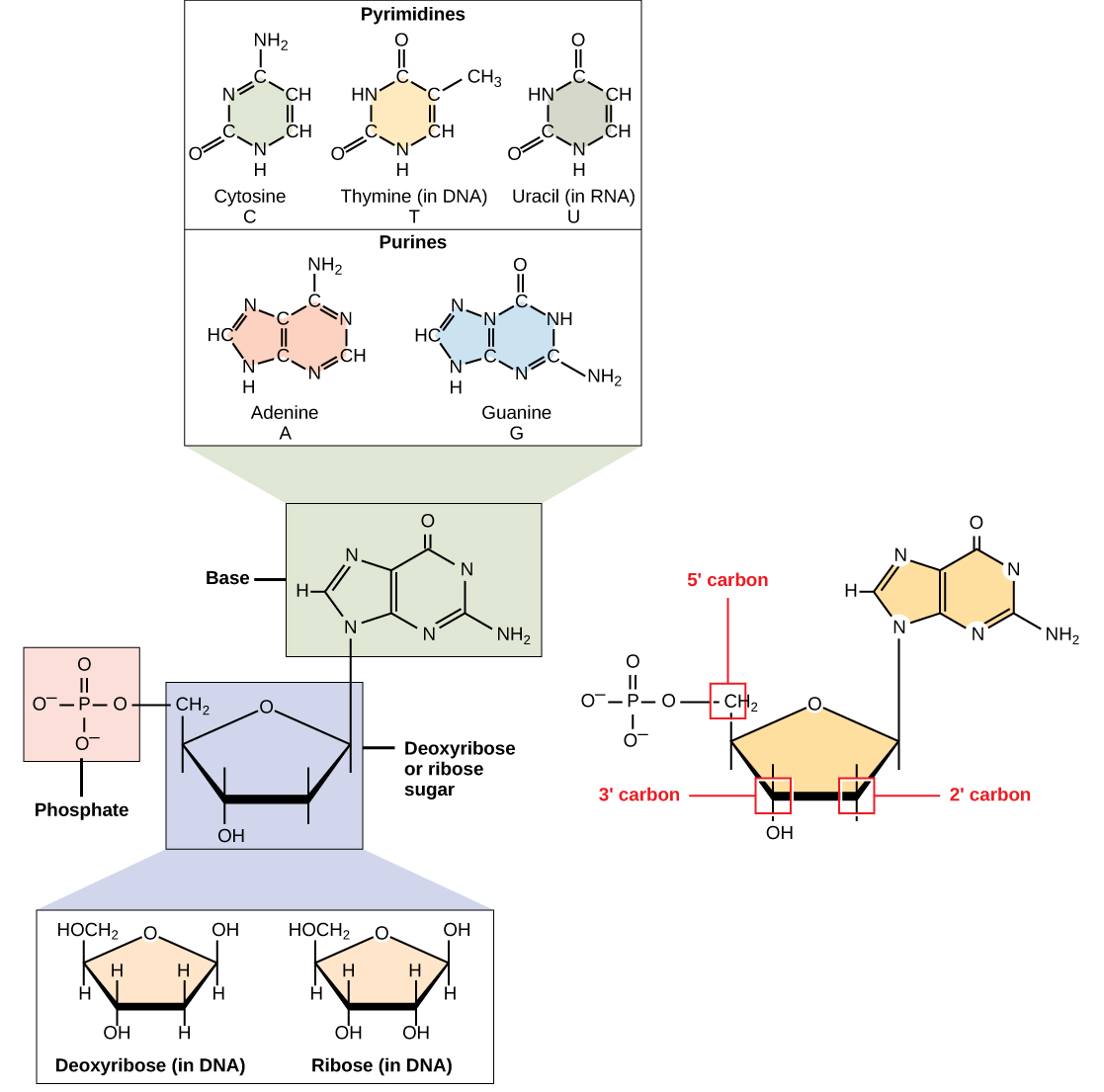
Die stickstoffhaltigen Basen, wichtige Bestandteile von Nukleotiden, sind organische Moleküle und werden so genannt, weil sie Kohlenstoff und Stickstoff enthalten. Sie sind Basen, weil sie eine Aminogruppe enthalten, die das Potenzial hat, einen zusätzlichen Wasserstoff zu binden, und somit die Wasserstoffionenkonzentration in ihrer Umgebung verringert, wodurch sie basischer wird. Jedes Nukleotid in DNA enthält eine von vier möglichen stickstoffhaltigen Basen: Adenin (A), Guanin (G) Cytosin (C) und Thymin (T).
Adenin und Guanin werden als Purine klassifiziert., Die Primärstruktur eines Purins besteht aus zwei Kohlenstoff-Stickstoff-Ringen. Cytosin, Thymin und Uracil werden als Pyrimidine klassifiziert, die einen einzigen Kohlenstoff-Stickstoff-Ring als Primärstruktur aufweisen (Abbildung \(\pageIndex{1}\)). Jeder dieser grundlegenden Kohlenstoff-Stickstoff-Ringe hat unterschiedliche funktionelle Gruppen. In der Molekularbiologie sind die stickstoffhaltigen Basen einfach durch ihre Symbole A, T, G, C und U bekannt.DNA enthält A, T, G und C, wohingegen RNA A, U, G und C enthält.
Der Pentosezucker in DNA ist Desoxyribose, und in RNA ist der Zucker Ribose (Abbildung \(\page Index{1}\)))., Der Unterschied zwischen den Zuckern besteht in der Anwesenheit der Hydroxylgruppe auf dem zweiten Kohlenstoff der Ribose und Wasserstoff auf dem zweiten Kohlenstoff der Desoxyribose (so dass Desoxyribose eine An-OH-Gruppe „fehlt“). Die Kohlenstoffatome des Zuckermoleküls sind nummeriert als 1′, 2′, 3′, 4′, und 5′ (1′ wird als „eine Primzahl“gelesen). Der Phosphatrest ist an die Hydroxylgruppe des 5′ Kohlenstoffs eines Zuckers und die Hydroxylgruppe des 3′ Kohlenstoffs des Zuckers des nächsten Nukleotids gebunden, die eine 5′-3′ Phosphodiesterverbindung bildet., Die Phosphodiesterverbindung wird nicht wie die anderen Verbindungen, die Monomere in Makromolekülen verbinden, durch einfache Dehydrationsreaktion gebildet: Ihre Bildung beinhaltet die Entfernung von zwei Phosphatgruppen. Ein Polynukleotid kann Tausende solcher Phosphodiesterverbindungen aufweisen.
Chargaff-Regeln
Als Watson und Crick in den 1940er Jahren die Struktur der DNA bestimmen wollten, war bereits bekannt, dass DNA aus einer Reihe besteht vier verschiedene Arten von Molekülen, Basen oder Nukleotide genannt: Adenin (A), Cytosin (C), Thymin (T), Guanin (G)., Watson und Crick wussten auch von Chargaffs Regeln, die eine Reihe von Beobachtungen über die relative Menge jedes Nukleotids waren, das in fast jedem DNA-Extrakt vorhanden war. Chargaff hatte beobachtet, dass für jede gegebene Spezies die Häufigkeit von A mit T identisch war und G mit C. Dies war für Watson & Cricks Modell unerlässlich.
Example \(\pageIndex{1}\)
Chargaff bestimmte die Zusammensetzung von Nukleinsäuren in Proben einer Vielzahl von Arten, einschließlich Prokaryoten und Eukaryoten. In einer Bakterienprobe betrug der Adeninanteil 15.,5% (Daten angepasst von Vscher et al., 1949). Welcher Anteil an Guanin wäre in dieser Stichprobe vorhanden gewesen und warum?
Lösung
Da Ein Paar mit T, sollte die Menge an T in etwa gleich A, oder etwa 15,5% Prozent. So, A + T = 15.5 + 15.5 = 31%.
der Prozentsatz Der G + C = 100% – 31% = 69%. Da G mit C paart, sollte die Menge von jedem von diesen ungefähr gleich sein, also jeweils ungefähr 34,5%.,
DNA-Doppelhelixstruktur
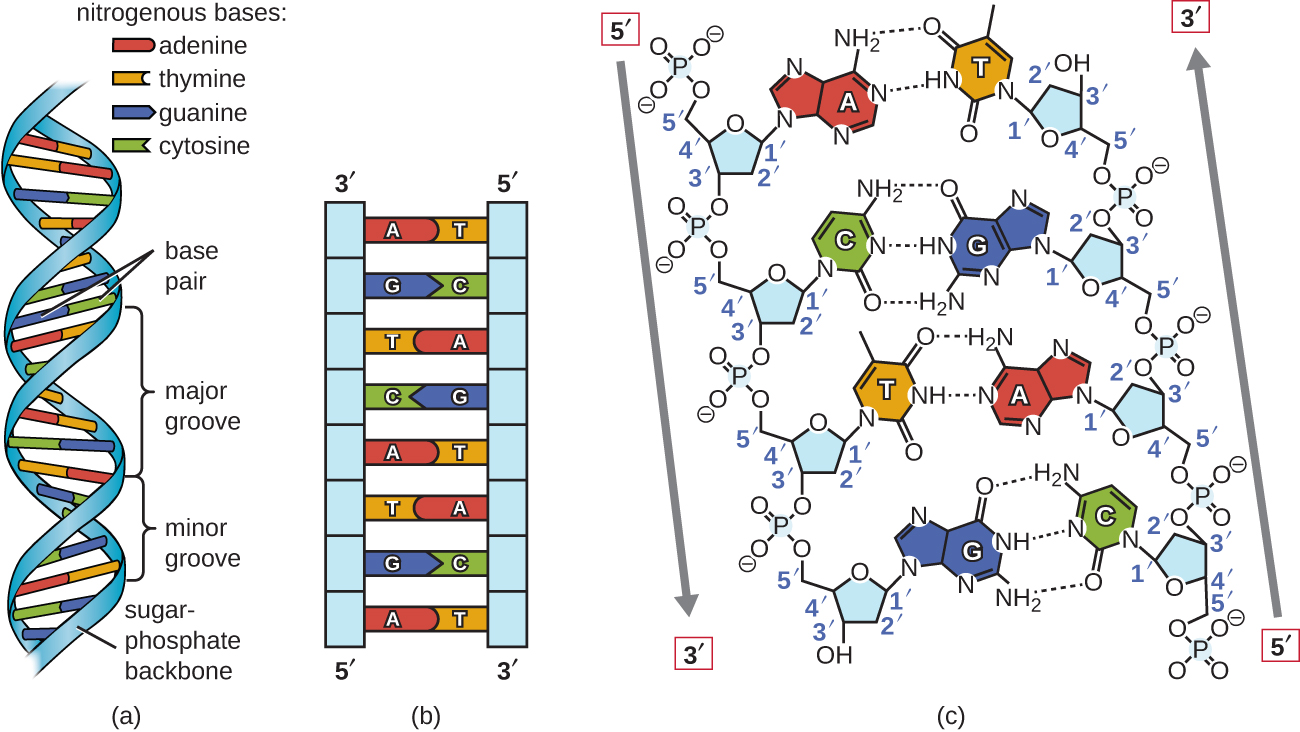
Unter Verwendung proportionaler Metallmodelle der einzelnen Nukleotide folgerten Watson und Crick eine Struktur für DNA, die mit Chargaffs Regeln und Röntgenkristallographiedaten übereinstimmte, die (mit einigen Kontroversen) von einem anderen Forscher namens Rosalind Franklin erhalten wurden. In Watson und Cricks berühmter Doppelhelix enthält jeder der beiden Stränge DNA-Basen, die durch kovalente Bindungen mit einem Zuckerphosphat-Rückgrat verbunden sind., Da eine Seite jedes Zuckermoleküls immer mit der gegenüberliegenden Seite des nächsten Zuckermoleküls verbunden ist, hat jeder DNA-Strang eine Polarität: Diese werden als 5′ (5-Primzahl) Ende und 3′ (3-Primzahl) Ende bezeichnet, entsprechend der Nomenklatur der Kohlenstoff in den Zuckern. Die beiden Stränge der Doppelhelix verlaufen in antiparallelen (d.h. entgegengesetzten) Richtungen, wobei das 5′ – Ende eines Strangs an das 3′ – Ende des anderen Strangs angrenzt. Die Doppelhelix hat eine Rechtshänderdrehung (anstelle der Linkshänderdrehung, die in populären Medien oft falsch dargestellt wird)., Die DNA-Basen erstrecken sich vom Rückgrat zum Zentrum der Helix, wobei ein Basenpaar aus jedem Strang Wasserstoffbrücken bildet, die helfen, die beiden Stränge zusammenzuhalten. Unter den meisten Bedingungen sind die beiden Stränge leicht versetzt, wodurch eine große Nut auf einer Seite der Doppelhelix und eine kleine Nut auf der anderen Seite entsteht. Aufgrund der Struktur der Basen kann A nur Wasserstoffbrücken mit T bilden, und G kann nur Wasserstoffbrücken mit C bilden (denken Sie an Chargaffs Regeln)., Jeder Strang soll daher komplementär zum anderen sein, und so enthält jeder Strang auch genügend Informationen, um als Vorlage für die Synthese des anderen zu fungieren. Diese komplementäre Redundanz ist wichtig für die DNA-Replikation und-Reparatur.
Nur bestimmte Arten der Basenpaarung sind erlaubt. Zum Beispiel kann ein bestimmtes Purin nur mit einem bestimmten Pyrimidin paaren. Dies bedeutet, dass A mit T und G mit C koppeln kann, wie in Abbildung \(\pageIndex{2}\) gezeigt. Dies wird als Basis-Komplementärregel bezeichnet. Mit anderen Worten, die DNA-Stränge sind komplementär zueinander., Wenn die Sequenz eines Strangs AATTGGCC ist, hätte der komplementäre Strang die Sequenz TTAACCGG. Während der DNA-Replikation wird jeder Strang kopiert, was zu einer Tochter-DNA-Doppelhelix führt, die einen elterlichen DNA-Strang und einen neu synthetisierten Strang enthält.
Thinking ahead exercise \(\paeindex{1}\)
Es tritt eine Mutation auf und Cytosin wird durch Adenin ersetzt. Welchen Einfluss wird dies Ihrer Meinung nach auf die DNA-Struktur haben?,
Antwort
Adenin ist größer als Cytosin und kann das Paar nicht richtig mit dem Guanin am gegenüberliegenden Strang basen. Dadurch wölbt sich die DNA. DNA-Reparaturenzyme können die Ausbuchtung erkennen und das falsche Nukleotid ersetzen.
Schreibe einen Kommentar